В предыдущих главах было показано, что флора и растительный покров тесно взаимосвязаны. Исследования методом конкретных флор в разных районах страны показали, что при широких географических сравнениях отдельные конкретные флоры целесообразно рассматривать как «точечные характеристики флористической ситуации (Юрцев, 1975, 1982; Галанин, 1987), так как при этом не столько важно, на какой площади они выявлены, а важно, чтобы они были полными и целостными в ландшафтном отношении, чтобы площадь их выявления охватывала все разнообразие биотопов элементарного флористического района в некоторой повторности (Толмачев, 1974). В разных природных зонах, на равнинах и в горах площади выявления конкретных флор колеблются в достаточно больших пределах - от 100 до 1000 км2 (Юрцев, 1975; Шмидт, 1980). Б. А. Юрцев (1987) считает, например, что разнообразие и площадь выявления конкретной флоры зависят от таких факторов, как история данного участка суши, длительность преемственного развития его биоты, степень контрастности ландшафтно-климатических условий минувших геологических эпох, интенсивность автохтонного видообразования, условия для сохранения реликтов и для иммиграции видов с соседних территорий.
Мы считаем, что конкретная флора, понимаемая как таксономическое разнообразие растительного покрова элементарного ландшафтного района, является его важнейшей характеристикой не только сама по себе как некоторое разнообразие, а как индикатор определенного комплекса эдафических, климатических, фитоценотических и исторических факторов, определяющих растительный покров этого района. Индикационная роль флоры традиционно используется при разделении растительного покрова на флористические царства и области при флористическом районировании (Тахтаджян, 1978).
Конкретные флоры - это не только точечные характеристики флористической ситуации. Они имеют вполне определенную внутриландшафтную эколого-ценотическую структуру (Галанин, 1973), которая зависит не только от характера рельефа и экологической структуры ландшафта, но и от исторических факторов. Именно здесь происходит стык традиционного флористического и геоботанического подходов к изучению растительного покрова, проводится отделение одной науки от другой, хотя, как показано нами в самом начале, флористика геоботаника отличаются не объектом, а предметом своего изучения. Объект изучения у геоботаники и флористики один, то растительный покров, а вот предмет изучения разный, у флористики - это таксономическое разнообразие растительного покрова, а у геоботаники - экологическое и эколого-ценотическое разнообразие.
Попробуем ответить на вопросы. В чем целостность конкретной флоры как системы? Кто должен изучать таксономический и ареалогический состав эколого-ценотических элементов - флорист или геоботаник? Как изменяются эколого-ценотические элементы конкретных флор в историческом плане? Эти и другие вопросы говорят о том, что проблема не столь проста, и окончательного ответа на многие вопросы еще не поручено. В последние десятилетия усиленно разрабатывались методы флористических исследований, развитие же теории флористики заметно отставало. Только в последние годы положение стало меняться, и даже центральные журналы стали отводить место дискуссии по теоретическим проблемам флористики и геоботаники. К сожалению, большим разнообразием подходов и точек зрения эта дискуссия пока не отличается. Именно историческая, а не только эколого-ценотическая обусловленность многих особенностей парциальной структуры конкретных флор привлекает к изучению «внутренней» структуры конкретных флор внимание ряда флористов (Галанин, 1973, 1974, 1987, 1990; Галанин, Марина, 1982; Марина, 1982, 1983, 1985; Юрцев, 1982, 1987; и др.). В этом случае флорист, как и геоботаник, имеет дело непосредственно с растительным покровом и всеми теми его категориями и подразделениями, которые отражают его внутриландшафтную дифференциацию. Представление о флоре как о множестве местных популяций видов соответствует современным представлениям о таксономической структуре растительного покрова. Недостаточно изучать флору как множество в эколого-ценотическом плане равноценных элементов безотносительно к объему их местных популяций, без учета характера взаимодействия этих популяций со средой обитания. При этом следует иметь в виду, что объем местной популяции вида характеризуется не только обилием этого вида в конкретных фитоценозах, но и широтой эколого-ценотической амплитуды вида в ландшафтном районе.
МЕТОДИКА
Важнейшей характеристикой флоры как дискриптивного множества несомненно являются видовой состав и таксономическая структура в целом (имеется в виду состав родов, семейств и других таксономических категорий). Вместе с тем на этом множестве могут быть выявлены и другие аспекты структуры: ареалогический, эколого-ценотический, спектр жизненных форм и т. д. Иными словами, на одном и том же множестве видов можно получить различные, разбиения на подмножества, получив при этом несколько аспектов структуры, или несколько фактор-множеств (Юрцев, Семкин, 1980; Галанин, Марина, 1982).
Обозначим фактор-множества А, В, ..... К, а их подмножества A1, А2, . . , Ai; B1, В2, . . . Вj; K1, K2, . . , Кe. Каждый вид конкретной флоры входит в одно из подмножеств каждого фактор-множества. В пределах одного фактор-множества подмножества не пересекаются, но подмножества, относящиеся к разным фактор-множествам, взаимно пересекаются, образуя матрицу отношений. Обозначим элементы пересечения двух фактор-множеств:
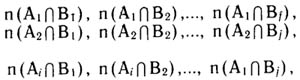
где  — число элементов (в нашем случае видов), одновременно принадлежащих подмножествам A1 и B1 фактор-множеств А и Б, и т. д.
— число элементов (в нашем случае видов), одновременно принадлежащих подмножествам A1 и B1 фактор-множеств А и Б, и т. д.
Система этих пересечений образует матрицу бинарных отношений, анализируя которую мы можем поставить вопрос о статистической независимости или, напротив, о сопряженности распределения видов по двум фактор-множествам (Галанин, 1974, 1987; Галанин, Марина, 1982). В случае отсутствия сопряженности ожидаемое число видов в подмножествах (элементах) пересечения можно рассчитать по формуле
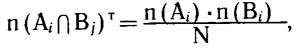
где n (Аi) - число видов в подмножестве Аi фактор-множества А; n(Вj) - число видов в подмножестве Бj фактор-множества В; N - общее число видов во флоре. При наличии сопряженности
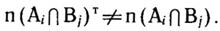
Статистическая достоверность сопряженности может быть оценена с помощью критерия Пирсона х2:
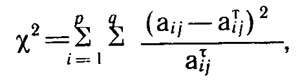
где q - число элементов в фактор-множестве А; Р - число элементов в фактор-множестве В;
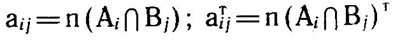
Расчитанное значение х2 сравнивается с табличным при k степенях свободы, где k = q(p - 1), и 5%-ном уровне значимости P0.05. Достоверность величины х2 оценивается как для матрицы в целом, так и отдельно для строк и столбцов и даже для отдельных элементов пересечения. В связи с тем что число подмножеств в фактор-множествах в разных конкретных флорах неодинаково, необходимо нормировать суммарное значение х2 матрицы числом элементов пересечения (число клеток матрицы бинарных отношений). Этот показатель т можно назвать «среднее значение х2 на одно подмножество пересечения»:
В принципе, сопряженный анализ фактор-множеств можно проводить не только с помощью матриц бинарных отношений, но и тринарных и т. д. В этом случае элементы пересечения будут тройными, четверными и т. д. Степень сопряженности при увеличении размерности матриц будет снижаться. Ясно, что для анализа сопряженности трех и более фактор-множеств конкретной флоры необходимо использовать компьютер. Ввод информации должен осуществляться путем перечисления видов конкретной флоры с указанием их принадлежности к определенным подмножествам вcех выделенных фактор-множеств. Настоящий метод сопряженного анализа структуры конкретных флор позволяет изучать флору естественной фитохории как некое закономерное целое, как систему видов и вышестоящих таксонов (Галанин, 1974, 1987, 1982, 1990). Эта система видов лимитирована разными факторами: разнообразием таксонов разного ранга, количеством и характером географических элементов, жизненных форм, эколого-ценотических элементов. Степень целостности флоры как системы видовых популяций невысока, это стохастическая система, но тем не менее географическое и экотопическое «маневрирование» популяций при перестройках флор, по-видимому, происходит сопряженно. К сожалению, конкретными и тем более достоверными данными по этому вопросу мы практически не располагаем.
Кроме флоры хребта Куркуре все виды изученных конкретных флор были разделены на два типа ареалогических (географических) элементов: широтные (А) - по протяженности географического ареала с севера на юг и долготные (В) - по протяженности географического ареала с запада на восток.
Оба фактор-множества и элементы их пересечения для каждой конкретной флоры приведены в таблицах. В каждом элементе пересечения приведено наблюдаемое число видов и ожидаемое при условии, что фактор-множества независимы и виды по элементам пересечения распределяются случайно. Значения х2 рассчитаны отдельно для широтных и долготных элементов и для конкретной флоры в целом.
Эколого-ценотические элементы конкретных флор выделялись по сходству их эколого-ценотических ареалов, т. е. по сходству распределения этих видов по типам и классам местообитаний в пределах каждого ландшафтного района. При выделении эколого-ценотических элементов использовались графы флористической структуры растительного покрова этих районов и распределения видов на этих графах. Примеры таких распределений и приведены в настоящей книге в предыдущих главах.
Эколого-ценотические элементы конкретной флоры также образуют фактор-множество. В свою очередь, это фактор-множество образует элементы пересечения с каждым из предыдущих фактор-множеств. Именно системы пересечения трех вышеуказанных фактор-множеств и составляют предмет анализа в настоящей главе.
ХАРАКТЕРИСТИКА РАЙОНОВ ИССЛЕДОВАНИЯ
Кроме флор районов, ландшафтно-ботаническая характеристика которых приведена в предыдущих главах настоящей монографии, при сопряженном анализе структуры флор использовались материалы по ряду других районов северной Евразии. Это район среднего течения р. Эргувеем на Чукотском полуострове, район среднего течения р. Цильмы - левого притока р. Печоры в Коми АССР, район окрестностей биостанции Сыктывкарского университета в среднем течении р. Вычегды в Коми АССР. Общая характеристика физико-географических условий, флоры и растительности этих районов приводится в настоящем разделе.
Среднее течение реки Эргувеем
Район Эргувеем расположен в пределах Охотско-Чукотского вулканогенного пояса. Сопочные массивы высотой 400-800 м над уровнем моря сочетаются здесь с обширными пространствами межгорных депрессий, в пределах которых наблюдаются всхолмления со щебнистыми бугорковатыми вершинами и интенсивно заболоченные, часто заполненные водой понижения. Склоны сопок террасированы, многие террасы перекрыты суглинком и заболочены. Плоские вершины сопок и нагорные террасы несут черты криогенной сортировки обломочного материала и имеют нанорельеф в виде систем каменных колец, опоясывающих бугорки.
Склоны сопок резко переходят в пологие шлейфы и конусы выноса, где поверхностный слой суглинка содержит включения грубого обломочного материала. Шлейфы и конусы выноса имеют грядовый и бугорковатый криогенный микрорельеф. Криогенные процессы наблюдаются и на склонах сопок средней крутизны (полосы, медальоны и т. д.), на крутых же склонах преобладают осыпи, в результате чего под такими склонами образуются россыпи незадернованного обломочного материала.
Для района характерна густая речная сеть. В долинах более крупных рек (Эргувеем, Ватамкайваам, Пепенвеем) хорошо выражены пойма и первые надпойменные террасы, в пределах которых можно выделить следующие элементы: галечные и песчаные пляжи, береговые валы, заболоченные участки первой надпойменной террасы, берега старичных озер, заиленные берега временных проток.
Климат района умеренно арктический, умеренно океанический (Клюкин, 1960). Средняя температура января приблизительно - 25°С, июля - +7°С, причем зимой возможны оттепели, а летом - заморозки и выпадение снега в любом месяце, садков за год выпадает около 700 мм, на холодное время приходится 120-130 мм. Лето короткое, снег стаивает в середине июня и выпадает во второй половине сентября. Благодаря расчлененному рельефу и сильным ветрам проводит перераспределение снега, и в депрессиях, оврагах, поймах рек и местах перегибов образуются огромные снежники, которые едва успевают полностью стаять к концу лета. На многих мелких речках и ручьях зимой образуются наледи площадью в несколько десятков гектаров.
Летняя облачность и большое количество жидких осадков способствуют переувлажнению почвы и заболачиванию. Вода часто сочится по шлейфам и широким ложбинам стока. Район расположен в зоне сплошного распространения вечной мерзлоты. Мерзлота, особенно в условиях влажного климата, является основным фактором формирования микро- и нанорельефа и обуславливает высокую подвижность субстрата даже на уровне мезорельефа.
Флора района Эргувеем насчитывает 297 видов сосудистых астений, из них арктических - 20%, аркто-альпийских - 52,5, гипоарктических и гипоарктомонтанных - 20,5, бореальных - 6, аркто-бореальных-1%. Долготный географический спектр флоры следующий, %: циркумполярных видов 35, широко распространенных в Евразии, не идущих в Америку - 8,7, узко распространенных в Евразии (к востоку от р. Колымы), не идущиx в Америку - 5, евразиатско-западноамериканских-16, собственно восточносибирско- и чукотско-западноамериканских (в широком смысле берингийских) - 12, сибирско- и восточно-сибирско-американских - 6, восточно-чукотско- американских - 0,3, узкораспространенных, ареал которых занимает Чукотский п-ов, п-ов Сьюард, острова Берингова и Чукотского морей (узкоберингийских) - 5. Приведенные цифры характеризуют флору как арктическую берингийского склада. Наиболее представительными по числу видов семействами являются злаки - 11, осоковые- 10, сложноцветные - 9, гвоздичные - 8, ивовые - 6, розоцветные - 6, камнеломковые - 6, норичниковые - 4, ситниковые - 4, бобовые - 3, верескоцветные -3. Спектр семейств также указывает на принадлежность нашей флоры к флорам берингийского сектора.
Исключительно широко в этом районе распространены сырые кочкарные осоково-пушицевые и осоково-кустарничковые тундры, которые занимают обширные участки надпойменных теppac, шлейфов склонов сопок, конуса выноса. В местах с интенсивным минеральным подтоком кочкарники обогащены видами разнотравья Saxifraga foliolosa, Senecio atripurpureus, S. frigidus, Pedicularis langsdorfii. На менее сырых участках тех же элементов рельефа встречаются сырые пятнистые осоковые тундры из Carex lugens с доминированием также Betula exilis, Salix pulchra, Vaccinium uliginosum. Среди них встречаются эвтрофные осочники и мохово-разнотравно-кустарничковые тундры. На щебнисто-суглинистых и сырых щебнистых субстратах тундры из Carex lugens сменяются разнотравно-осоково-кустарничковыми сырыми пятнистыми тундрами, в которых виды сухих местообитаний (Antennaria friesiana, Охуtropis czukotica, Artemisia furcata, A. borealis и др.) сочетаются с видами влаголюбивыми (Juncus biglumis, J. albescens, Beckwithia chamissonis, Carex misandra). Это, вероятно, связано с непостоянством степени увлажненности местообитаний в течение вегетационного периода. В начале лета мерзлый слой почвы лежит очень близко к поверхности и, являясь водоупором, приводит к переувлажнению корнеобитаемого горизонта почвы. Во второй половине лета глубина протаивания резко увеличивается и корнеобитаемый горизонт почвы хорошо дренируется.
Кассиопейные тундры благодаря большей снежности местообитаний распространены на Чукотском полуострове гораздо шире, чем на Западной Чукотке: на Раучуа и в Чаунской низменности. Возле снежников встречаются нивальные тундры специфичным набором видов: Salix chamissonis, S. polaris, Poa paucispicula, Oxyria digyna, Nardosmia glacialis, Phippsia algida и др. Там, где снег стаивает до начала июля, эти тундры переходят в нивально-луговинные с Artemisia arctica, Potentilla emarginata, Thalictrum alpinum и др. На пологих щебнистых склонах, выпуклых частях нагорных террас распространены пятнистые кустарничково-лишайниковые тундры: кассиопейные, дриадовые, диапенсиевые, часто доминантой является и Salix phlebophylla.
Растительность склонов сопок на Эргувеем слабо изменяется с их экспозицией, что можно объяснить преобладай рассеянной радиации. На отдельных участках встречаются эвтрофные тундры, в составе которых довольно много видов представителей восточночукотского кальцифильного комплекса Potentilla biflora, Artemisia globularis, Drias integrifolia, Senecio kjellmanii, Anemone parviflora, Puccinellia wrightii и др.
Поймы рек несут отчетливые черты нивальности. Специфичные пойменные виды приурочены к сухим песчаным и галечным наносам, образуя своеобразные луговины. В поймах встречаются низкорослые ивняки до 1,5 м высотой из Salix alaxensis, S. pulchra, S. hastata.
В прибрежно-водной растительности доминируют Carex stans, Eriophorum russeolum, E. polystachyon, E. medium. Среди прибрежно-водных видов есть арктические: Ranunculus pallasii, Dupontia psilosantha, Pleuropogon sabinii, Arctophila fulva и бореальные: Hippuris vulgaris, Ranunculus reptans, Sparganium hyperboreum и др.
В целом растительность района среднего течения р. Эргувеем, расположенного южнее районов среднего течения р. Раучуа и устья р. Чаун, но более подверженного воздействию холодных морских ветров, несет гораздо больше арктических черт, что оправдывает отнесение района к подзоне северных гипоарктических тундр (Юрцев, 1973). Более детальная характеристика флоры и растительности этого района дана в наших публикациях (Галанин, 1973, 1974, 1989).
Среднее течение реки Цильмы
Бассейн р. Цильмы - левого притока р. Печоры - расположен на северо-западе Коми АССР. Верхнее и среднее течение этой реки проходит по Тиманскому кряжу. В геоботаническом отношении район исследования относится к подзоне крайне северной тайги Тимано-Печорской подпровинции, Ижемскому геоботаническому округу сосновых лесов (Юдин, 1978).
Рельеф района равнинный или полого-увалистый, сильнее изрезанный вблизи рек. Поверхность сложена мощной толщей флювиогляциальных песчаных отложений, перекрывающих более ранние глинистые четвертичные отложения, которые местами выходят на поверхность (Юдин, 1978). Четвертичные глинистые отложения содержат мало обменных оснований, кислые, что способствует развитию подзолообразования. На междуречьях встречаются обширные депрессии, в большинстве случаев занятые болотами, в основном верхового и переходного типов. На долю таких болот в районе приходится до 32% площади (Панов и др., 1980). На боровых террасах преобладают песчаные железистые подзолистые и подзолисто-глеевые почвы, на междуречьях - подзолисто-глеевые суглинистые и супесчаные, в депрессиях - торфянисто-глеевые и торфяные (Забоева, 1975).
Современный рельеф сформировался в плейстоцене и голоцене. Район покрывался ледниками в днепровскую, московскую и валдайские ледниковые эпохи, а в конце ледниковых эпох частично, вероятно, затоплялся водами морских трансгрессий (Былинский, 1970). Наиболее возвышенные части Тиманского кряжа во время трансгрессий не погружались, но, по всей вероятности, были покрыты в это время льдом.
Низовья Цильмы относятся к умеренно-холодному агроклиматическому району. Осадков за год выпадает около 500 мм, безморозный период длится в среднем 80 дней, период же с устойчивым снежным покровом длится 180 дней. Средняя высота снежного покрова 81 см, средняя температура января -28°С, июля 14,5°С (Агроклиматические ресурсы..., 1973).
В районе широко распространены зеленомошные и лишайниковые сосновые леса. Вдали от рек на внутренних частях боровых террас преобладают заболоченные зеленомошно-сфагновые кустарничковые сосновые леса и сфагновые олиготрофные болота. Еловых лесов немного, они встречаются на суглинистых междуречьях и в долинах рек на надпойменных террасах. Для сосновых и еловых древостоев характерна постоянная примесь лиственницы. Тундровые элементы растительности в виде сообществ с доминированием карликовой березки встречаются изредка в долинах небольших речек. В пойме Цильмы на значительных площадях распространены луга несомненно антропогенного происхождения. Эти луга возникли в результате расчистки пойменных ивняков, сведения пойменных травяных и зеленомошных ельников.
Флора района, по нашим данным, насчитывает 327 видов и рас сосудистых растений. Ведущими по числу видов являются следующие семейства: сложноцветные (10,4%), злаковые (9,8), осоковые (9,1), розоцветные (6,7), лютиковые (5,5), гвоздичные (4,0), крестоцветные (4,0), ивовые (3,6), гречишные (3,3), норичниковые (2,9%). Евразиатских видов во флоре 30%, циркумполярных и почти циркумполярных - 35, европейских- 16, азиатских - 7%. По широтным ареалогическим группам виды флоры распределяются следующим образом: бореальных - 63%, плюризональных-12, гипоарктических и аркто-альпийских - 9%.
Полевые исследования на Цильме проводились летом 1980 г. Более детальная характеристика флоры и растительности, а также флористической структуры растительного покрова этого района дана в специальной работе (Галанин, Носов, 1989).
Окрестности биостанции Сыктывкарского университета в среднем течении реки Вычегды
Биостанция Сыктывкарского университета расположена в 120 км восточнее г. Сыктывкара на правом берегу Вычегды в урочище «Белый яр». Климат района умеренно континентальный с длительной холодной зимой и коротким умеренно теплым и влажным летом. Возможная продолжительность солнечного сияния составляет 4535 ч/году, но частая облачность сокращает ее почти в 3 раза. Радиационный баланс небольшой - 30 ккал/см2 в год. Преобладает западно-восточный перенос воздушных масс умеренных широт, довольно часто происходит заток арктического воздуха с Ледовитого океана и редко проявляются прорывы тропического воздуха.
Годовая сумма осадков 670 мм, из них 350 мм испаряется с поверхности. В среднем к 22 апреля разрушается устойчивый снеговой покров. Полностью же он стаивает в начале мая. В это время освобождается от льда Вычегда и среднесуточная температура воздуха достигает 5°С. Вегетация растений начинается в начале мая. Весной часты заморозки, во время которых температура иногда снижается до -15°С. Летние дни очень длинные, солнце не заходит по 18 ч/сут. В отдельные дни температура повышается до 30 и даже 35°С, но по мере вторжения арктического воздуха она в течение суток может снизиться на 20-25°. Для района исследования сумма температур воздуха выше 10° составляет в среднем 1477°, а сумма температур выше 15° составляет в среднем 763°. Пасмурных дней летом мало, обложные дожди наблюдаются редко. Более характерны ливневые осадки, во время которых выпадает до 30-40 мм осадков за один дождь. В конце августа появляются первые признаки осени. Среднесуточная температура воздуха становится ниже 5° в конце сентября. В это время заканчивается вегетационный период. Средняя температура января -15,1°С, а июля 16,6°С. При составлении климатической характеристики наряду с собственными данными использованы данные из «Агроклиматического справочника Коми АССР» (1973).
Русло Вычегды преимущественно песчаное и сильно подвержено деформации. Большая часть ландшафтного района приходится на долину реки. Обширная пойменная терраса высотой 2-4 м изрезана многочисленными старицами, протоками и озерами, лежащими между песчаных гряд. Пойменная часть долины ежегодно затапливается, пик половодья в среднем наблюдается 16 мая, средняя продолжительность половодья около 50 дней. Вода во время половодья поднимается до отметки 5 м над нулем графика. Спад половодья происходит более замедленно, чем подъем, летней межени река достигает в конце июня-середине июля, при этом уровень воды в среднем понижается до 33 см ниже нуля графика. Осенью наблюдается 2-3 дождевых паводка, высота подъема которых колеблется от 0,5 до 2,5 м. К затопляемым элементам долины в окрестностях биостанции относятся боровые террасы и отдельные наиболее возвышенные гряды в пойме.
Все озера района пойменного происхождения, они в той или иной мере связаны с рекой. Вскрытие озер ото льда отстает от вскрытия на реке на 1-2 дня, а замерзание происходит на 2 недели раньше. В летнее время озера у поверхности прогреваются до 26°С, на глубине же температура бывает на 10° и более ниже, чем у поверхности. Помимо озер, большое количество осадков задерживают болота.
Почвенный покров окрестностей биостанции характерен для подзоны типичных подзолистых почв средней тайги. Здесь можно наблюдать болотные почвы низинного, переходного и верхового типов. Наиболее же широко распространены аллювиальные почвы пойменных террас. Основная почвообразующая порода - супесь или песок, подстилаемый завалуненным моренным суглинком. Боровые террасы сложены песками. Для таежных участков водоразделов характерны типичные сильно подзолистые почвы с почти полным отсутствием дернового процесса. Развиваются они в хорошо дренированных условиях при глубоком залегании грунтовых вод. Песчаные почвы боровых террас под борами беломошными относятся к железистым подзолам. Под смешанными лесами развиты гумусово-железистые подзолы. Весьма широко в окрестностях биостанции распространены болотно-подзолистые почвы. На первой стадии заболачивания лесов формируются торфянисто-подзолисто-глеевые почвы. На участках с большим увлажнением они переходят в торфяно-подзолисто-глеевые почвы. Для лугов характерны пойменные дерновые почвы, местами с признаками оглеения. Дерновые почвы поймы имеют наиболее благоприятное сочетание химических и физических свойств.
В растительном покрове окрестностей биостанции можно выделить следующие наиболее контрастные сообщества растений: прибрежно-водная растительность озер и стариц, заболоченные осоковые и осоково-разнотравные пойменные луга низкого уровня, злаково-осоково-разнотравные пойменные луга среднего уровня, сухие злаково-разнотравные пойменные луга высокого уровня на песчаных грядах, сухие боры беломошники, сосняки зеленомошно-черничные, сосняки заболоченные сфагново-кустарничковые, сфагновые олиготрофные болота на вогнутых участках надпойменной террасы, смешанные сосново-елово-пихтовые разнотравно-кустарниковые леса на участках высокой поймы, сосново-мелколиственные разнотравно-кустарничковые леса на участках высокой поймы, оригинальные антропогенные группировки на катищах - местах сосредоточения заготовленного леса перед его сплавом.
Участки низкой поймы испытывают наиболее мощное аллювиальное воздействие. Здесь широко распространены разнотравные ивняки, перемежающиеся небольшими по площади прогалинами с луговой разнотравно-злаковой растительностью.
Следует отметить, что пойменные луга имеют здесь антропогенное происхождение. На участках поймы, заливаемых паводковыми водами на длительный период, эти сообщества вполне устойчивы и только через 20-30 лет, при снятии антропогенного фактора, постепенно сменяются ивняками и пойменными смешанными лесами. Смена луга лесом ускоряется при ослаблении аллювиального режима. Однако слишком интенсивный аллювиальный процесс приводит к разрушению луговой растительности, размыву дернины и формированию песчаных пляжей.
Варьирование видового состава лугов и изменение обилия видов при переходе от одного участка к другому связано с изменением рельефа и ценотическими факторами. В процессе конкуренции на каждом участке растительного покрова формируется группировка видов и жизненных форм, оптимальная или почти оптимальная в данных эдафических условиях и в рамках данной конкретной флоры.
Инвентаризация флоры в окрестностях биостанции осуществлялась во время полевых практик по ботанике в 1980 и 1981 гг. В ней принимали активное участие студенты и лаборанты кафедры ботаники Сыктывкарского университета С. В. Дегтева, О. В. Жданова, а также коллеги зоологи Г. Н. Романов, В. Н. Гурьев и В. А. Соловьев. Всем им выражаю признательность.
РЕЗУЛЬТАТЫ СОПРЯЖЕННОГО АНАЛИЗА СТРУКТУРЫ КОНКРЕТНЫХ ФЛОР
Использование метода сопряженного анализа типологических ареалогических и эколого-ценотических структур конкретных флор позволяет получить дополнительную информацию об их генезисе (Галанин, 1974, 1987; Галанин, Марина, 1982), показывает, что разные эколого-ценотические элементы конкретных пор имеют достоверную отличающуюся ареалогическую структуру.
Флора хребта Куркуре
Ареалогическому и генетическому анализу флоры горного Алтая посвящена обширная литература (Крылов, 1981; 1931; Ревердатто, 1934, 1947; 1965; Ильин, 1941, 1958; Альбицкая, 1946; Куминова, 1946, 1963; Соболевская, 1946, 1958; Калинина, 1948; Шретер, 1953; Положий, 1964, 1972; Малышев, 1968; Положий, Крапивка, 1971, 1978; Пленник, 1976; Краснобородов, 1977; и др.). История флоры и растительности этого региона в общих чертах установлена, выявлены основные этапы формирования флоры, характер и роль миграционных потоков, роль автохтонного видообразования.
В настоящем разделе рассмотрим особенности генезиса растительного покрова хребта Куркуре в послеледниковую эпоху на фоне общих закономерностей, установленных предыдущими исследователями. При изучении географического распространения видов флоры хребта Куркуре были использованы различные флористические сводки (Крылов, 1927-1964; Черепнин, 1957-1967; «Арктическая флора СССР», 1960- 1984; «Растения...», 1963-1977; Малышев, 1965, 1968; Егорова, 1968; Скворцов, 1968; Коропачинский, 1975; Оразова, 1975; Красноборов, 1976; Цвелев, 1975; «Ареалы...», 1977; Еленевский, 1978; Седельников, 1979; Липшиц, 1979; Ревушкин, 1988). Проведенный анализ и сравнение позволили выявить следующие типы географических ареалов, охватывающие в своей совокупности все разнообразие географического распространения видов флоры Восточного Алтая.
1. Циркумполярные и почти циркумполярные плюризональные и бореальные виды, включая и виды космополиты: Equisetum pratensis, Androsace septentrionalis, Gentiana tenella, Hieracium umbellatum, Huperzia selago, Vaccinium vitis-idaea, Linnaea borealis, Moneses uniflora, Fragaria vesca, Rosa acicularis, Lamium album, Equisetum palustre, Caltha palustris, Hippuris vulgaris и др. Всего 88 видов.
2. Циркумполярные, почти циркумполярные и евразиатские аркто-альпийские виды: Woodsia glabella, Saxifraga cernua, Hierochloe alpina, Poa alpina, Luzula confusa, L. parviflora, Cardamine bellidifolia, Oxyria digyna, Minuartia biflora, Thalictrum alpinum, Carex atrofusca, Potentilla nivea, Salix glauca, Cystopteris dickieana, Juncus triglumis и др. Всего 80 видов.
3. Евразиатские плюризональные и евразиатские бореальные виды: Carex globularis, С. laxa, Cypripedium guttatum, Populus tremula, Betula humilis, Paeonia anomala, Salix lapponum, Isoetes echinospora, Trisetum sibiricum, Festuca valesiaca, Lilium martagon, Polygonum bistorta, Solidago virgaurea, Antennaria dioica, Chelidonium majus, Spiraea media, Bupleurum aureum и др. Всего 82 вида.
4. Азиатские, преимущественно сибирские плюризональные и бореальные виды: Rubus sachalinensis, Poa sibirica, P. ursulensis, Potentilla sericea, Zygadenus sibiricus, Stellaria bungeana, Cerastium pauciflorum, Lonicera pallasii, Salix divaricata, Ribes altissima, Iris ruthenica, Trollius asiaticus, Pedicularis compacta, Pinus sibirica, Abies sibirica, Larix sibirica и др. Всего 76 видов.
5. Центральноазиатские виды: Carex orbicularis, Sedum ewersii, Dryadanthe tetrandra, Dracocephalum grandiflorum, D. imberbe, Galium saurense, Leontopodium ochroleucum, Taraxacum brevirostre, Euphrasia regelii, Lonicera hispida и др. Всего 11 видов.
6. Южносибирско-среднеазиатские виды, ареал которых включает горы Южной Сибири от Забайкалья до Средней (иногда Передней) Азии; заходят в северную часть Монголии: Juniperus pseudosabina, Poa tianschanica, Carex melanocephala, C. dichroa, Primula pallasii, Calamagrostis pavlovii, Schulzia crinita, Draba oreades, Rhodiola algida, R. coccinea, Trollius lilacinus, Lagotis integrifolia, Doronicum altaicum, Corydalis pauciflora и др. Всего 45 видов.
7. Южносибирско-северомонгольские виды. Ареалы включают горы юга Сибири и Северную Монголию, нередко простираются до Саура, Тарбагатая и Джунгарского Алатау: Роа krylovii, Allium amphibolum, A. nutans, Berberis sibirica, Pedicularis elata, Festuca tristis, Rheum altaicum, Aquilegia sibirica, Bergenia crassifolia, Rosa oxyacantha, Scrophularia altaica, Libanotis monstrosa, Salix turczaninovii, Macropodium nivale, Saxifraga terektensis и др. Всего 57 видов.
8. Алтайские эндемики и гемиэндемики: Senecio vereczaginii, Potentilla elegans, P. gelida, Draba dasycarpa, Dendranthe-ma sinuatum, Aconitum decipiens, Alchemilla altaica, Thymus schischkinii, Euphrasia krylovii, Cirsium komarovii и др. Всего 10 видов.
Перечисленная совокупность типов географических ареалов характеризует географическую структуру данной конкретной флоры, которая, с одной стороны, определяется особенностями генезиса растительного покрова данного ландшафтного района, т. е. флористическими связями его с соседними территориями, а с другой - набором местообитаний, динамикой климата и ценотическими взаимодействиями видов. Становление и динамика флоры зависят от факторов экологических, климатических и ценотических. Например, на первых этапах заселения освобождающейся из-под ледника территории основную роль играют факторы флористические (наличие подходящих для поселения видов в ближайших рефугиумах), экологические и климатические. На последующих этапах, по мере приближения растительности к климаксовому состоянию, все большее значение в формировании флоры играют факторы ценотические.
Таксономическая неоднородность растительного покрова в пределах ландшафтного района должна рассматриваться в двух аспектах - экологическом и историческом. Миграционно-активными являются не все виды локальной флоры, а лишь те, для которых в районе имеются подходящие местообитания, лежащие в пределах их эколого-ценотического оптимума. В нашем случае в качестве возможных доноров выступали прителецкие и тувинско-монгольские рефугиумы. Различные местообитания хребта Куркуре «брали» из рефугиумов разные виды, одни брали виды таежного фитоценотического комплекса, другие - степного и лесостепного (имеется в виду комплекс видов лиственничной экспозиционной лесостепи), третьи - альпийского и горнотундрового. В субнивальном и верхней части альпийского пояса флористические изменения в плейстоцене, вероятно, были незначительными, флора здесь развивалась преемственно.
Для более детального ареалогического анализа истории формирования растительного покрова Восточного Алтая была выявлена экотопологическая структура флоры хребта Куркуре (Галанин, 1979). На основе сходства распределений видов на графе флористической структуры растительного покрова флора была разделена на несколько крупных эколого-ценотических комплексов.
1. Растения лесов субальпийского и верхней части лесного поясов: Lycopodium annotinum, Chamaenerion angustifolium, Rubus saxatilis, Equisetum sylvaticum, Cypripedium guttatum, Populus tremula, Corydalis capnoides, Pulmonaria molissima, Trientalis europaea, Rubus sachalinensis, Zygadenus sibiricus, Calamagrostis pavlovii и др. Всего в этот комплекс отнесено 57 видов.
2. Эвритопные виды субальпийского пояса: Equisetum arvense, Сагех media, С. ledebouriana, Anthoxanthum odoratum, Juniperus sibiricus, Pachypleurum alpinum, Silene chamarensis, Luzula sibirica и др. Всего 14 видов.
3. Виды субальпийских лугов. В эту группу включены растения, имеющие высокую активность на субальпийских лугах. В других сообществах субальпийского и верхней части лесного поясов они встречаются, но довольно спорадично и при этом приурочены к травянистым фитоценозам или их фрагментам: Gentiana tenella, Erigeron elongatus, Synchaeta norvegica, Aconitum napellus, Thalictrum minus, Helictotrichon pubescens, Trisetum sibiricum, Veratrum lobelianum, Allium sibiricum, Dianthus superbus, Cerastium pseudosibiricum, Veronica longifolia, Polygonum bistorta, Delphinium elatum, Trifolium lupinaster, Pedicularis compacta и др. Всего 152 вида.
4. Растения пойменных кустарников субальпийского пояса: Pentaphylloides fruticosa, Chamaenerion latifolium, Astragalus frigidus, Equisetum hyemale, Carex caespitosa, C. eleusinoides, Salix lapponum, S. hastata, S. glauca, S. divaricata, S. krylovii, Rhodiola rosea, Ribes nigrum и др. Всего 33 вида.
5. Растения заболоченных местообитаний субальпийского пояса: Calamagrostis lapponica, Eriophorum polystachyon, Baeothryon alpinum, B. caespitosum, Carex magellanica ssp. irrigua, C. rostrata, C. dioica, C. mollissima, Juncus filiformis, Rubus chamaemorus, Epilobium palustre, Ledum palustre, Vaccinium uliginosum, Oxycoccus microcarpus, Galium uliginosum, Salix caesia и др. Всего 39 видов.
6. Виды водных местообитаний: Isoetes lacustris, I. echinospora, Sparganium minimum, Potamogeton graminifolius, P. perfoliatus, Batrachium divaricatum, Callitriche palustris, Myriophyllum spicatum, Hippuris vulgaris, Ranunculus natans и др. Всего 13 видов.
7. Растения скал и осыпей: Polypodium vulgare, Campanula rotundifolia, Woodsia glabella, W. ilvensis, Dryopteris fragrans, Cryptogramma stelleri, Draba fladnizensis, D. lanceolata, D. oreades, Saxifraga cernua, S. sibirica, Potentilla ornithopoda, Chelidonium majus, Cystopteris fragilis, Salix vestita, S. berberifolia, Thesium repens, Pseudostellaria rupestris, Orostachys spinosa, Potentilla desertorum, Rhodiola coccinea, Rheum altaicum и др. Всего 47 видов.
8. Виды, встречающиеся преимущественно в тундроподобных сообществах: Equisetum variegatum, Huperzia selago, Coeloglossum viride, Pyrola rotundifolia, Linnaea borealis, Diphasium alpinum, Anemone narcissiflora var. crinita, Eutrema edwardsii, Dryas oxyodonta, Thermopsis alpina и др. Всего 17 видов.
9. Растения альпийских лугов: Oxyria digyna, Sibbaldia procumbens, Carex tripartita, Pedicularis amoena, Patentilla gelida, Primula nivalis, Ranunculus nivalis, R. altaicus, Taraxacum brevirostre, Paracolpodium altaicum, Trollius lilacinus, Veronica densifolia, Lagotis integrifolia, Doronicum altaicum и др. Всего 59 видов.
10. Высокоальпийские виды. Встречаются преимущественно в субнивальном поясе: Hierochloe alpina, Poa alpina, P. altaica, Luzula confusa, L. spicata, Lloydia serotina, Minuartia arctica, Cardamine bellidifolia, Carex rupestris, Saxifraga flagellaris ssp. setigera, S. terektensis, Senecio resedifolius, Dryadanthe tetrandra, Papaver canescens и др. Всего 18 видов.
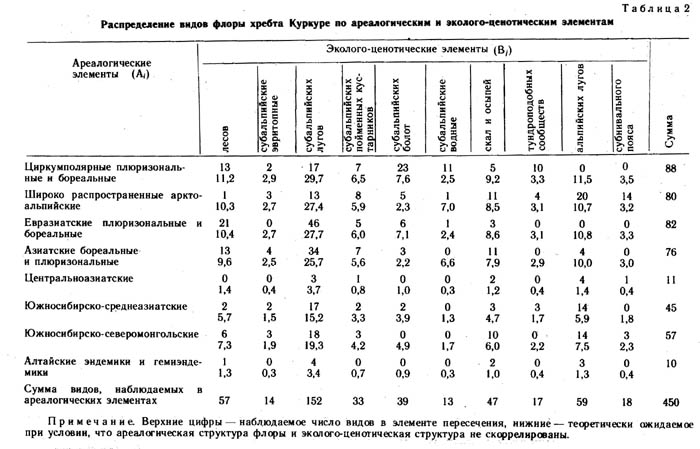
Перечисленными эколого-ценотическими комплексами в общих чертах характеризуется экотопологическая структура флоры хребта Куркуре. На корреляцию между географическим распространением видов и их распределением по местообитаниям указывает ряд авторов (Юрцев, 1968; Галанин, 1973, 1987; Красноборов, 1976). Для более достоверного анализа скоррелированности ареалогической и эколого-ценотической структур флоры нами использован метод Пирсона. Составлена таблица соотносительного распределения видов флоры по ареалогическим и эколого-ценотическим элементам (табл. 2). Элементы пересечения двух фактор-множеств заполнены эмпирическими числами видов (верхняя цифра) и рассчитанными на основании краевых сумм ожидаемыми числами видов при условии, что ареалогическая и эколого-ценотическая структуры флоры не скоррелированы (нижние цифры). В табл. 3 приведены отклонения, оцененные с помощью критерия Пирсона для всех строк, столбцов, и суммарное значение для всей таблицы в целом. Пороговые значения критерия х2 при 95%-ном уровне значимости в нашем случае составляют: при одной степени свободы (для одной отдельной клетки) х2 =3,84, при 9 степенях свободы (для отдельной строки) х2=16,9 при 7 (для отдельного столбца) х2 =14,1, для таблицы в целом х2 = 113,2. Наряду с величинами отклонений для каждого элемента таблицы приводится знак отклонения (табл. 3).
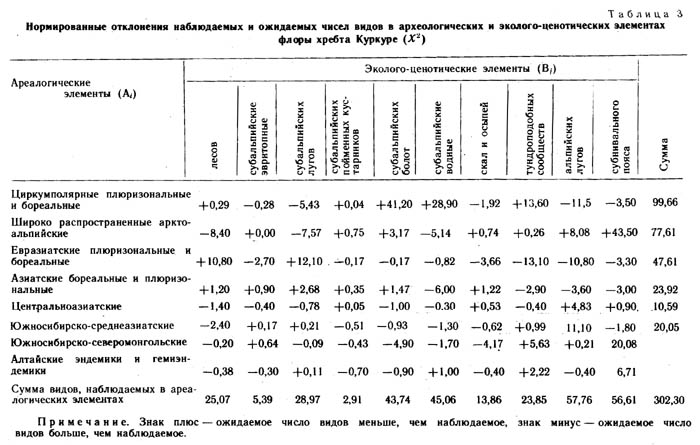
Полученные результаты показали, что между эколого-ценотической и ареалогической структурами флоры хребта Куркуре имеет место достоверная сопряженность. Так, виды с циркумполярными плюризональными и бореальными ареалами положительно сопряжены с заболоченными, водными и тундровыми местообитаниями, отрицательно же - с субальпийскими и альпийскими лугами. Этот факт можно объяснить тем, что флористический состав болотных, водных и тундровых фитоценозов наименее изменчив во времени, эти сообщества более стабильны и не только флористически, но, по-видимому, и ценотически и биогеохимически. Напротив, луговые фитоценозы менее стабильны и в них видов с обширными ареалами меньше, чем следует из случайной модели распределения.
Широко распространенные арктоальпийские виды положительно сопряжены с местообитаниями субнивального пояса и альпийскими лугами, отрицательно - с лесами и субальпийскими лугами. Данный факт также вполне соответствует нашим представлениям о характере аркто-альпийских видов, их экологии и географии. Несколько неожиданно недостаточное количество аркто-альпийских видов на субальпийских лугах. Евразиатские плюризональные и бореальные виды положительно сопряжены с лесами и субальпийскими лугами, отрицательно- с альпийскими лугами, скалами и осыпями. Этот факт также не требует специального объяснения, хотя «избыток» видов данной группы на субальпийских лугах довольно неожиданный. Азиатские (сибирские) бореальные и плюризональные виды отрицательно сопряжены с водными местообитаниями. Южносибирско-среднеазиатские положительно сопряжены с альпийскими лугами, отрицательной же достоверной связи не обнаруживают ни с какими эколого-ценотическими комплексами. Это значит, что флора альпийских лугов Восточного Алтая в значительной степени формировалась за счет иммиграции видов с гор Средней Азии.
Южносибирско-северомонгольские виды положительно сопряжены с альпийскими лугами и отрицательно со скалами и осыпями, а также с заболоченными местообитаниями. Весьма интересно, что роль южносибирско-северомонгольского региона в формировании флоры альпийских лугов значительна. По всей вероятности, такое видообразование активизировалось здесь в гумидные эпохи, несомненно совпадающие с периодами оледенений. Центральноазиатские виды не имеют достоверного отрицательного сопряжения, но положительно сопряжены с альпийскими лугами. Алтайские эндемики и гемиэндемики недостоверно положительно сопряжены с альпийскими лугами. Все эти сопряженности позволяют предполагать, что гумидный период в геологической и экологической истории Южной Сибири и Центральной Азии был довольно продолжительным и стабильным.
Рассмотрим характер сопряжения эколого-ценотических элементов с ареалогическими элементами. Так, комплекс видов лесов положительно сопряжен с евразиатскими плюризональными и бореальными видами и отрицательно - с аркто-альпийскими. Несколько неожиданно отрицательное сопряжение комплекса видов эвритопных субальпийского пояса с аркто-альпийской группой. Комплекс видов субальпийских лугов положительно сопряжен с евразиатской плюризональной и бореальной группой, но отрицательно с циркумполярной плюризональной и аркто-альпийской группами. По всей вероятности, флора субальпийских лугов Восточного Алтая начала формироваться еще в ледниковую эпоху ниже ледника за счет видов лесного пояса. Леса в то время, вероятно, почти вплотную подходили к окончаниям долинных и каровых ледников, где перемежались с прогалинами, занятыми травянистой растительностью лугового типа. Приток альпийских и аркто-альпийских видов на эти лесные приледниковые луга был затруднен, и немалую роль в создании этого барьера играли ценотические факторы.
Комплекс видов пойменных кустарников субальпийского пояса не имеет достоверного сопряжения ни с одной из ареалогических групп видов. На заболоченных местообитаниях наборы видов формировались за счет циркумполярных, плюризональных бореальных видов, вероятно из прителецкого рефугиума. Комплекс растений скал и осыпей отрицательно сопряжен с южно-сибирско-северомонгольским ареалогическим элементом. Тундры хребта Куркуре формировались за счет циркумполярных плюризональных и бореальных видов, флористически они очень слабо обособлены от других местообитаний данного района. По всей вероятности, это связано с молодостью тундрового типа растительности на Восточном Алтае и затруднительным процессом миграции тундровых видов с соседних горных систем Сибири. Можно предположить даже отсутствие свободного флористического обмена между тундрами Восточного Алтая Арктики. Ерниковые сообщества в ледниковую эпоху на восточном Алтае были распространены незначительно, и по краю ледника, видимо, росли темнохвойные и светлохвойные леса, перемежающиеся, как было отмечено выше, лесными лугами. Такие луга в верхней части лесного пояса здесь встречаются поныне.
Становление ерникового пояса происходило по мере отступания ледника и аридизации климата. В современную эпоху связи с некоторой гумидизацией климата мохово-лишайниковые ерники стали замещаться ерниками мохово-разнотравными луговой растительностью. Так, А. В. Куминова (1963) считает, что изменение растительного покрова Алтая в современную эпоху связано с гумидизацией климата, так что субальпийские луга и высокогорные тундры - явление более молодое, чем альпийские луга. Наши данные подтверждают го предположение.
Для альпийских лугов и местообитаний субнивального пояса характерно положительное сопряжение с аркто-альпийским, южносибирско-среднеазиатским, южносибирско-северомонгольским и центральноазиатским ареалогическими элементами при отрицательном сопряжении с циркумполярными, евразийскими и азиатскими (сибирскими) плюризональными и бореальными элементами. Это все указывает на относительную древность флористического комплекса видов альпийского и субнивального поясов, так как значительное число видов формировалось в довольно узком регионе и эти виды можно считать эндемами.
Сопряженный анализ флоры хребта Куркуре оставляет впечатление как бы двойственности ее видового состава. Экологические условия альпийского и субнивального поясов в горах таковы, что приспособление (имеются в виду морфофизиологические адаптации) к ним выходцев из нижних поясов затруднено. Нельзя не согласиться с мнением А. И. Толмачева (1958) о том, что приспособление растений к определенным термическим условиям, отличным от тех, в которых складывались приспособительные черты строения их предков, совершается в естественных условиях лишь с большим трудом и весьма медленно. Таксономический вакуум при формировании растительности высокогорий заполняется не за счет автохтонного видообразования, а за счет иммигрантов с соседних гор. Таким образом, флора альпийского и субнивального поясов очень консервативна.
Уже в конце третичного периода на Алтае в горах в виде самостоятельного пояса была представлена темнохвойная тайга (Куминова, 1963) и альпийский пояс был отделен ею от «весенней» флоры эфемеров широколиственных лесов и черневой тайги. Считается, что именно некоторые представители весенней флоры листопадных широколиственных лесов, по всей вероятности, были предшественниками флоры альпийского пояса (Толмачев, 1958). В широколиственных лесах эти виды прошли предадаптацию к короткому вегетационному периоду, выработали способность большую часть года находиться в состоянии корневищ и луковиц. В отличие от растений, обитающих под пологом темнохвойной тайги, эфемеры и эфемероиды широколиственных лесов приспособлены к яркому освещению, что также могло пригодиться обитателям субальпийских и альпийских лугов.
Формирование флоры альпийского и субнивального поясов на Алтае шло преимущественно миграционным путем. Темнохвойная тайга ничего не могла «предложить» альпийскому поясу, а черневая тайга и широколиственные леса, богатые весенними эфемерами, оставались в предгорьях. Автохтонное видообразование имело место в горах Южной Сибири и Северной Монголии, узкие эндемы формировались и в альпийском поясе Алтая, и даже Восточного Алтая, но это автохтонное видообразование, по-видимому, шло на базе альпийских видов, мигрировавших сюда через горы Средней Азии с Кавказа и Гималаев.
В связи с общим похолоданием климата и усилением континентальности в середине плейстоцена темнохвойная тайга стала сменяться светлохвойной лиственничной. Формация лиственничных лесов оказалась более преемственной по отношению к представителям неморальной весенней флоры, чем формация темнохвойной тайги. Лиственничная тайга и высокогорные степи смогли «питать» процессы альпийского видообразования, и эти процессы во второй половине плейстоцена оживились, особенно на Юго-Восточном Алтае.
По всей вероятности, плейстоценовое оледенение Восточного Алтая не было покровным, при этом выше ледников не только сохранялись растительные сообщества, но могли осуществляться миграции видов на большие расстояния путем обмена видами между отдельными вершинами и горными цепями, возвышавшимися над ледником. Вообще следует признать, что широтные миграции в истории растительного покрова и флор Голарктики мели большое значение во все геологические эпохи, а не только в ледниковую (Толмачев, 1958). Каналом для миграции видов при этом служили горные цепи Евразии и Америки.
Таким образом, флора хребта Куркуре в плане генезиса может быть разделена на две части. Флора субнивального и альпийского поясов является весьма устойчивым и древним образованием. Она не уничтожалась в эпохи плейстоценовых оледенений и развивалась непрерывно, по крайней мере на протяжении плейстоцена. Напротив, флора субальпийского пояса возникла на базе флоры лесного пояса после отступления ледника. Сразу вслед за ледником, отступавшим в результате таяния, освободившиеся экотопы оказались занятыми высокотравными лугами, ивняками и ерниковыми тундроподобными сообществами. По мере формирования почвенного покрова и в связи с некоторой гумидизацией климата мохово-лишайниковые ерники замещались ерниками мохово-разнотравными и высокотравными лугами.
Решающую роль в становлении флоры хребта Куркуре несомненно играл прителецкий рефугиум. В эпоху оледенения, одним из центров которого был хребет Куркуре, долина р. Чулышман (во всяком случае ниже устья р. Шавлы) и Телецкого озера была свободна от льда. Лишь отдельные ледниковые языки «выдавливались» с Чулышманского плоскогорья и свисали в Чулышманское ущелье. В устьях рек Чульчи, Шавлы и особенно Кыги в эпоху оледенения сохранялись участки черневой тайги с комплексом третичных реликтов. Основу флоры рефугиумов составляли виды таежной генетической группы (Куминова, 1963): Pinus sibirica, Abies sibirica, Picea obovata, Majanthemum bifolium, Oxalis acetosella, Linnaea borealis, Paris quadrifolia, Orthilia secunda, Pyrola rotundifolia, Vaccinium vitis-idaea, V. myrtillus, Lycopodium annotinum, Carex globularis и др. Однако наряду с ними встречались и представители неморальной группы. Так, в нижней части лесного пояса в долине р. Кыги, впадающей в Телецкое озеро в его южной части, обнаружены Galium krylovii, G. paradoxum, Brunnera sibirica, Dryopteris filix-mas, Festuca gigantea, F. altissima, Asarum europaeum, Asperula odorata, Poa remota, Daphne mezereum, Carpesium triste, Polystichum braunii, Epilobium montanum, Cardamine impatiens, Actaea erytrocarpa, A. spicata, Carex sylvatica (Галанин и др., 1977; Блузманас и др., 1979).
Тувинско-монгольский рефугиум в становлении растительного покрова исследуемого района в конце плейстоцена - начале голоцена заметной роли, вероятно, не играл. Однако многие виды горной лесной формации, проникшие сюда в ксеротермическую эпоху межледниковья, ледниковый период могли пережить в долине Чулышмана.
Флора районов Эргувеем и Раучуа (сравнительный анализ)
При типизации географических ареалов флор районов Эргувеем и Раучуа использована информация, заимствованная из источников, перечисленных в начале предыдущего раздела, а также из «Флоры Аляски и близлежащих территорий» (Hulten, 1968), «Флоры СССР» и некоторых других источников. Для характеристики ареалов видов в данном случае нами принято деление северных и умеренно-северных областей земного шара на три широтных пояса, которые соответствуют крупным широтным поясам растительности.
1. Арктический (острова и побережье морей Ледовитого океана, занятые арктическими пустынями и тундрами).
2. Гипоарктический (северная часть таежной зоны, лесотундру и южную часть тундровой зоны).
3. Бореальный (зона тайги, включая полосу хвойно-широколиственных лесов).
В соответствии с этими поясами все виды разделены на несколько широтных ареалогических групп: арктическую, гипоарктическую и бореальную. Арктические и гипоарктические группы ареалов, в свою очередь, подразделены на подгруппы: собственно арктические виды, аркто-альпийские (последние кроме Арктики распространены за ее пределами в южных районах, но здесь всегда встречаются только в высокогорьях), собственно гипоарктические виды, гипоаркто-монтанные виды (от предыдущих отличаются тем, что могут встречаться южнее гипоарктики, но только в горах, где явно тяготеют к верхним поясам).
Наряду с широтными группами ареалов выделялись долготные группы по степени простирания ареалов с запада на восток, т. е. в меридиональном направлении. В связи с этим северное полушарие разделено нами на следующие долготные сектора:
1. Европейский (Европа до Урала, а также Кавказ и Закавказье).
2. Сибирский (часть Северной Азии от Урала до Чукотского полуострова включительно).
3. Восточносибирский (часть предыдущего сектора от Енисея до Берингова пролива).
4. Собственно восточносибирский (часть предыдущего сектора от Лены до Берингова пролива).
5. Чукотский (часть предыдущего сектора от Колымы до Берингова пролива).
6. Восточночукотский (часть предыдущего сектора от Ам-гуэмы до Берингова пролива).
7. Западноамериканский (часть Северной Америки от Берингова пролива до Макензи).
8. Американский (включает предыдущий сектор и простирается на всю Северную Америку до побережья Атлантики).
9. Евразиатский (от Атлантического побережья Евразии до Берингова пролива).
10. Сибирско-американский (территория от Урала к востоку до Атлантического побережья Северной Америки).
11. Сибирско-западноамериканский (от Урала на восток до Макензи).
12. Восточносибирско-американский (от Енисея на восток до Атлантического побережья Северной Америки).
13. Восточносибирско-западноамериканский (от Енисея на восток до Макензи). Это в широком смысле Берингийский сектор или Мегаберингия по Б.А. Юрцеву (1974).
14. Собственно восточносибирско-западноамерйканский (от Лены до Макензи).
15. Чукотско-американский (от Колымы до Атлантического побережья Северной Америки).
16. Чукотско-западноамериканский (от Колымы на восток до Макензи. Этот сектор можно назвать собственно Берингийским).
17. Восточночукотско-американский (от Амгуэмы на восток до Атлантического побережья Северной Америки).
18. Восточночукотско-крайнезападноамериканский (от Амгуэмы до континентальных районов Аляски. Этот сектор можно назвать узко Берингийским).
19. Евро-американо-восточносибирский (Европа, Северная Америка и восточная часть северной Азии).
20. Циркумполярный (простирается на все континенты вокруг северного полюса).
При выделении долготных ареалогических элементов флоры мы следовали принципу увеличения детализации по направлению к району исследования. Следует отметить, что выделение групп ареалов несколько условно, так как двух совершенно сходных географических ареалов просто не существует. Мы имеем дело с типичным ареалогическим континуумом, который условно разрезаем на удобные для нас части, чтобы решить вполне определенные задачи ареалогического анализа.
К группе арктических видов отнесены Deschampsia brevifolia, Pleuropogon sabinii, Dupontia psilosantha, Phippsia algida, Puccinellia wrighfii, Festuca hyperborea, F. brevissima, Eriophorum callitrix, Carex rariflora, Salix arctica, S. reptans, Rumex beringensis, R. arcticus, Sagina intermedia, Minuartia rubella, Ranunculus pallasii, Cardamine digitata, Cochlearia arctica, Saxifraga eschscholtzii, S. grandipetala, Chrysosplenium wrightii, Armeria arctica, Pedicularis pennellii и др.
К группе аркто-альпийских видов отнесены Woodsia glabella, Cystopteris dickieana, Dryopteris fragrans, Hierochloe alpina, Alopecurus alpinus, Trisetum spicatum, Poa malacantha, Poa alpina, P. alpigena, P. paucispicula, Festuca brachyphylla, Carex rupestris, C. atrofusca, C. misandra, C. holostoma, C. podocarpa, Juncus biglumis, Luzula rufescens, Tofieldia coccinea, Lloydia serotina, Salix reticulata, S. polaris, Oxyria digyna, Oxygraphis glacialis, Ranunculus nivalis, R. sulphureus, Cassiope tetragona и др.
Группа гипоарктических видов включает Agrostis borealis, Calamagrostis lapponica, Poa sublanata, Poa glauca, Arctophila fulva, Festuca altaica, Roegneria borealis, Elymus interior, Hordeum jubatum, Eriophorum russeolum, Carex lugens, C. melanocarpa, C. algida, C. rotundata, Veratrum oxysepalum, Salix glauca, S. pulchra, Betula exilis, Cerastium maximum, Arenaria tschuktschorum, Ranunculus grayi, Pedicularis labradorica, Empetrum nigrum и др.
К бореальным видам отнесены Equisetum limosum, Botrychium lunaria, Alopecurus aequalis, Agrostis trinii, Calamagrostis purpurea, Poa pratensis, Festuca rubra, Bromus sibiricus, Roegneria confusa, R. macroura, Eriophorum polystachyon, Carex chordorrhiza, C. aquatilis, C. rostrata, Smilacina trifolia, Chosenia arbutifolia, Salix myrtilloides, Stellaria palustris, Pulsatilla multifida, Ranunculus reptans, Parnassia palustris, Comarum palustre и др.
Долготные ареалогические группы можно проиллюстрировать ареалами следующих конкретных видов. Так, в группе циркумполярных и почти циркумполярных рассматриваются Woodsia ilvensis, Dryopteris fragrans, Hierochloe alpina, Deschampsia brevifolia, Pleuropogon sabinii, Poa arctica, P. pratensis, P. alpigena, P. glauca, Dupontia psilosantha, Festuca rubra, F. brachyphylla, Eriophorum russeolum, Kobresia bellardii, Carex rupestris, C. chordorrhiza, C. tenuiflora, C. tripartita, C. rariflora, Juncus biglumis, Luzula confusa, L. nivalis, Salix glauca, Oxyria digyna, Koenigia islandica, Polygonum viviparum, Sagina intermedia, Ranunculus pallasii, R. reptans, Cardamine bellidifolia, Saxifraga cernua, S. hirculus и др.
Евразиатско-западноамериканские: Calamagrostis holmii, Carex obtusata, C. williamsii, C. saxatilis, Luzula tundricola, Lloydia serotina, Salix polaris, S. hastata, Rumex graminilolius, R. arcticus, Cerastium maximum, Minuartia arctica, Dianthus repens, Parrya nudicaulis, Alyssum biovulatum, Rubus arcticus, Dryas octopetala, D. punctata, Hedysarum obscurum, Polemonium boreale, P. acutiflorum, Myosotis asiatica, Pedicularis verticillata и др.
Сибирско-американские: Potamogeton subsibiricum, Hierochloe pauciflora, Festuca auriculata, Bromus pumpellianus, Eriophorum callitrix, Carex algida, C. fuscidula, Tofieldia coccinea, Salix pulchra, Polygonum humifusum, Caltha arctica, Pulsatilla multifida, Ranunculus affinis, Novosieversia glacialis, Ledum decumbens и др.
Берингийские или восточносибирско-западноамериканские: Selaginella sibirica, Arctagrostis arundinacea, Agrostis trinii, Poa malacantha, P. paucispicula, Puccinellia wrightii, Hordeum jubatum, Carex bonanzensis, C. eleusinoides, C. podocarpa, C. tolmiei, Luzula rufescens, L. beringensis, Veratrum oxysepalum, Salix phlebophylla, S. sphenophylla, S. fuscescens, S. alaxensis, Betula exilis, Polygonum ellipticum, Stellaria jumbellata, Arenaria capillaris, Beckwithia chamissonis, Papaver macounii, Cardamine digitata, Corydalis arctica, Rhodiola atropurpurea, Saxifraga firma, S. eschscholtzii и др.
Американские, заходящие на восток Северной Азии: Carex lugens, С. supina ssp. spaniocarpa, C. membranacea, Smilacina trifolia, Silene acaulis, Anemone richardsonii, A. parviflora, Ranunculus grayi, Erysimum pallasii, Chrysosplenium tetrandrum, Parnassia kotzebuei, Dryas integrifolia, Oxytropis maydelliana, Arctous erytrocarpa, Pedicularis langsdorfii, P. sudetica ssp. albolabiata, P. capitata, Saussurea angustifolia, Crepis nana и др.
Евразиатские и сибирские, не заходящие в Америку: Poa sublanata, Carex melanocarpa, С. pediformis, C. ledebouriana, Luzula parviflora, Allium strictum, Salix nummularia, S. reptans, S. myrtilloides, Polygonum laxmanii, Stellaria palustris, Minuartia verna, Caltha caespitosa, Oxygraphis glacialis, Chrysosplenium alternifolium, Dryas ineisa, Astragalus frigidus, A. schelechovii, A. kolymensis, Oxytropis middendorffii, Pachypleurum alpinum, Phlojodicarpus villosus, Eritrichium villosum, Pedicularis amoena, P. sceptrum-carolinum, Crepis chrysantha и др.
Восточносибирские и чукотские, не заходящие в Америку: Calamagrostis angustifolia, Helictotrichon dahuricum, Roegneria confusa, R. jacutensis, Carex appendiculata, Scirpus maximowiczii, Chosenia arbutifolia, Salix tschuktschorum, S. saxatilis, S. kolymensis, S. krylovii, Rumex beringensis, Polygonum tripterocarpum, Claytonia acutifolia, C. arctica, Arenaria tschuktschorum, Silene stenophylla, Delphinium chamissonis, Pulsatilla dahurica, Papaver microcarpum ssp. czekanowskii, Cardamine victoris, Oxytropis vassilczenkoi, Rhododendron parvifolium, Dracocephalum palmatum, Castilleja hyparctica и др.
 |
Всего в обоих районах нами обнаружено 417 видов и рас сосудистых растений, из них 212 встречаются в обоих районах (общие для Раучуа и Эргувеем), 108 видов встречаются в районе Раучуа и не встречаются на Эргувеем, 85 видов встречаются в районе Эргувеем и не встречаются на Раучуа. Коэффициент сходства флор, по Жаккару, 52%.
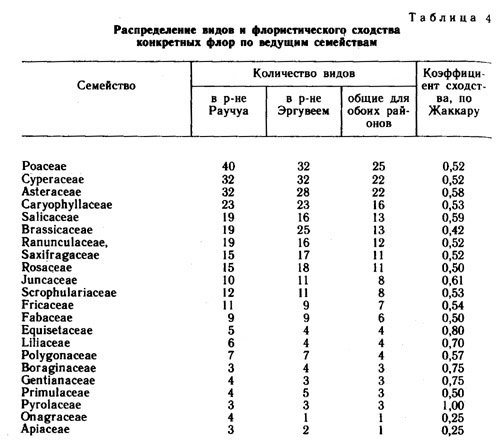
В табл. 4 приведен список наиболее представительных по числу видов семейств. Для каждого из этих семейств подсчитаны числа видов во флоре Эргувеем, флоре Раучуа и в совокупности видов, общих для обоих районов. Кроме того, в таблице приведены коэффициенты флористического сходства, по Жаккару, рассчитанные отдельно по каждому семейству. Из 10 ведущих семейств наши флоры наиболее сходны по наборам видов семейств ивовых и сложноцветных, а наименее - по наборам видов крестоцветных. Интересно, что наиболее высокие коэффициенты сходства приходятся на малочисленные семейства - хвощевые, лилейные, бурачниковые, горечавковые и первоцветные. В наибольшей же степени рассматриваемые флоры отличаются семействами кипрейных и зонтичных. По всей вероятности, наибольшее сходство между флорами должно обнаруживаться в тех семействах, где процессы видообразования идут медленнее и где виды имеют более широкие эколого-ценотические амплитуды. |
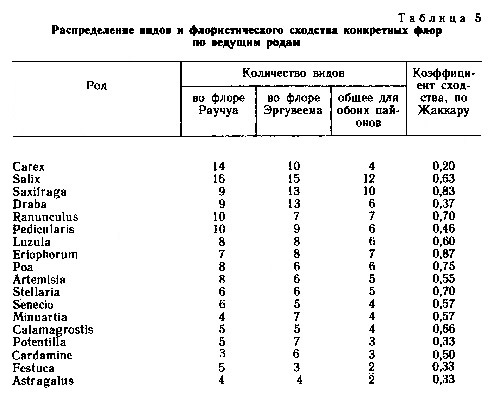
Наиболее представительными родами (табл. 5) во флорах Эргувеем и Раучуа являются ивы, осоки, камнеломки, крупки, лютики, мытники, ожики, пушицы, мятлики и полыни. Наиболее сходны сравниваемые флоры по наборам видов родов камнеломки, лютики, пушицы и звездчатки, представители которых, видимо, менее чувствительны к градиенту климата, разделяющему Восточную и Западную Чукотку. Наименьшее сходство между флорами приходится на род осок, что несомненно связано с климатическими различиями районов и тем, что виды в роде осок экологически более дифференцированы по градиенту увлажнения. Чувствительны к градиенту континентальности - океаничности климата виды в родах крупка, лапчатка, овсяница и астрагал.
Порядок расположения в ранжированных рядах ведущих семейств и ведущих родов в обоих флорах весьма сходный. Это показывает, что история формирования сравниваемых флор в значительной степени была сходной.
В результате сравнения внутриландшафтных эколого-ценотических ареалов видов во флоре района среднего течения р. Эргувеем было выделено семь эколого-ценотических элементов. |
|
1. Виды с очень широкой эколого-ценотической амплитудой, встречающиеся в самых разных местообитаниях - от равнинных кочкарных осоково-пушицево-кустарничковых тундр до щебнисто-суглинистых и щебнистых пятнистых разнотравно-кустарничковых мохово-лишайниковых и лишайниковых тундр. Это Hierochloe alpina, Carex lugens, Salix pulchra, Betula exilis, Polygonum viviparum, Cassiope tetragona, Ledum decumbens, Vaccinium uliginosum, V. vitis-idaea, Diapensia obovata, Dryas punctata, Luzula confusa, Eriophorum vaginatum, E. polystachyon, Empetrum nigrum и др.
2. Виды сухих горных тундр. Этот элемент объединяет виды, встречающиеся во всех вариантах щебнистых тундр, для которых характерен недостаток почвенной влаги на большей части вегетационного периода: Rhododendron kamtschaticum, Saxifraga firma, Artemisia furcata, Oxytropis czukotica, Novo-sieversia glacialis, Potentilla elegans, Silene stenophylla, Potentilla uniflora, Antennaria friesiana, Saxifraga eschscholtzii, Draba stenopetala, Ermania parryoides, Dryas incanescens, Oxytropis mertensiana, Claytonia arctica и др.
3. Виды равнинных осоково-пушицевых кустарничковых тундр: Carex stans, Salix fuscescens, Carex algida, С. rotundata, Pedicularis pennellii, Carex misandra, Arctagrostis latifolia, Pedicularis sudetica ssp. albolebiata, Saxifraga foliolosa, Juncus biglumis, Carex membranacea, Luzula tundricola, Rubus chamaemorus, Polygonum ellipticum, Juncus castaneus и др.
4. Виды сырых евтрофных тундр: Salix reticulata, Juncus triglumis, Beckwithia chamissonis, Lagotis minor, Tofieldia coccinea, Thalictrum alpinum, Claytonia acutifolia, Carex atrofusca, C. fuscidula, C. williamsii, Saxifraga hirculus, S. hieracifolia, Gentiana algida, Senecio frigidus, Carex holostoma, Rumex arcticus, Luzula nivalis, Saussurea angustifolia, Rhododendron parvifolium и др.
5. Виды нивальных луговин: Oxyria digyna, Trisetum spicatum, Salix polaris, Artemisia arctica, Saxifraga hyperborea, Cardarnine bellidifolia, Gentiana glauca, Poa paucispicula, Nardosmia frigida, N. glacialis, Taraxacum alascanum, Anemone sibirica, Phippsia algida, Carex podocarpa, Anemone richardsonii и др.
6. Виды пойменных ивняков и пойменных редкотравных лугов: Festuca altaica, Potentilla emarginata, Chamaenerion latifolium, Androsace ochotense, Elymus interior, Carex eleusinoides, Veratrum oxysepalum, Merckia physodes, Cardarnine pratensis, Astragalus alpinus, A. frigidus, Galium boreale, Aster sibiricus, Taraxacum ceratophorum, Salix alaxensis, S. hastata, S. krylovii, Calamagrostis purpurea, Bromus pumpellianus, Salix nummularia, S. lanata и др.
7. Виды прибрежно-водных местообитаний: Potamogeton subsibiricum, Sparganium hyperboreum, Arctophila fulva, Ranunculus pallasii, R. reptans, R. gmelinii, Hippuris vulgaris, Caltha arctica, Eriophorum scheuchzeri, Cardamine victoris, Pleuropogon sabinii, Dupontia psilosantha, Hierochloe pauciflora и др.
Во флоре района среднего течения р. Раучуа по сходству эколого-ценотических ареалов выделено восемь эколого-ценотических элементов.
1. Виды с очень широкой эколого-ценотической амплитудой: Dryas punctata, Empetrum nigrum, Vaccinium uliginosum, V. vitis-idaea, Ledum decumbens, Betula exilis, Arctagrostis latifolia, Carex lugens, Salix pulchra, Empetrum nigrum, Arctous alpina, Polygonum tripterocarpum и др.
2. Виды сухих щебнистых тундр: Hierochloe alpina, Artemisia arctica, Saxifraga firma, Anemone sibirica, Luzula confusa, Poa glauca, Arenaria tschuktschorum, Dracocephalum palmatum, Dianthus repens, Erysimum pallasii, Pulsatilla multifida, Festuca auriculata, Potentilla nivea, Festuca brachyphylla, Salix phlebophylla, Silene stenophylla, Oxytropis czukotica, Polygonum laxmanii, Carex pediformis, Papaver czekanowskii, Artemisia furcata, Selaginella sibirica и др.
3. Виды сырых евтрофных тундр: Claytonia acutifolia, Parrya nudicaulis, Luzula nivalis, Lagotis minor, Salix reticulata, Polygonum viviparum, Carex misandra, C. algida, Pedicularis oederi, Ramischia obtusata, Senecio frigidus, Rhododendron parvifolium, Thalictrum alpinum, Corydalis arctica, Carex williamsii, C. fuscidula, Tofieldia coccinea, Saxifraga hirculus.
4. Виды пойменных ивняков и лугов: Calamagrostis purpurea, Chamaenerion latifolium, Galium boreale, Poa pratense, Tanacetum boreale, Equisetum arvense, Artemisia kruhsiana, Festuca altaica, Roegneria borealis, R. jacutensis, Agrostis trinii, Artemisia vulgaris, Elymus interior, Festuca rubra, F. cryophila, Trisetum molle, Allium schoenoprasum, Carex eleusinoides, Salix alaxensis, S. kolymensis, S. hastata, Artemisia tilesii, Salix schwerinii, Botrychium lunaria, Adoxa moschatellina, Chosenia arbutifolia и др.
5. Виды нивальных луговин: Nardosmia glacialis, Salix polaris, Huperzia selago, Salix polaris, Oxyria digyna, Ranunculus pygmaeus, Saxifraga porsildiana, S. tenuis, S. hyperborea, Potentilla elegans, Poa paucispicula, Carex tripartita и др.
6. Виды прибрежно-водных местообитаний: Equisetum limosum, Sparganium hyperboreum, Alopecurus aequalis, Carex aquatilis, C. rostrata, C. saxatilis, Ranunculus pallasii, R. reptans, R. gmelinii, Hippuris vulgaris, Menyanthes trifoliata, Comarum palustre и др.
Эколого-ценотическая активность видов в сравниваемых районах отличается весьма существенно. Наиболее сильно она различается у видов, приведенных ниже:
Виды, более активные на Эргувеем:
Poa mafacantha, Eriophorum vaginatum, E. polystachyon, Carex stans, Salix phlebophylla, S. fuscescens, Polygonum viviparum, Valeriana capitata, Artemisia arctica, Poa glauca, Poa arctica, Veratrum oxysepalum, Stellaria ciliatosepala, Anemone sibirica, Minuartia arctica, M. macrocarpa, Cassiope tetragona, Diapensia obovata, Carex misandra, C. rotundata, C. algida, Juncus biglumis, J. castaneus, Salix reticulata.
Виды, более активные на Раучуа:
Saxifraga foliolosa, Novosieversia glacialis, Potentilla elegans, Oxytropis czukotica, Pedicularis pennellii, Artemisia furcata, Antennaria friesiana, Festuca brachyphylla, Pedicularis lapponica, Pyrola incarnata, Nardosmia frigida, Calamagrostis holmii, C. purpurea, Equisetum arvense, Galium boreale, Potentilla nivea, Selaginella sibirica, Pedicularis amoena, Senecio tundricola, Cardamine bellidifolia, Calamagrostis lapponica, Salix saxatilis, Rubus chamaemorus, Erysimum pallasii.
Ареалогический географический состав этих двух групп видов приведен ниже (первая цифра - виды, более активные на Эргувеем; вторая - виды, более активные на Раучуа).
Циркумполярные -11; 11
Берингийские – 8; 3
Сибирские, не идущие в Америку – 1; 3
Американские, идущие в Азию – 1; 1
Сибирские, идущие в Америку – 3; 2
Сибирско-американские – 1; 1
Евразиатские, идущие в Америку - 0; 2
Аркто-альпийские - 17; 9
Арктические – 3; 1
Аркто-бореальные – 2; 0
Гипоарктические 3; 5
Гипоаркто-монтанные – 0; 3
Бореальные – 0; 3
Для сравнительного анализа двух конкретных флор по соотношению в них разных географических элементов мы выделили пять совокупностей видов: 1 - виды, общие для обоих районов, встречающиеся как на Раучуа, так и на Эргувееме; 2 - виды, специфичные для района Раучуа, встречающиеся в этом районе и не встречающиеся на Эргувееме; 3 - виды, специфичные для района Эргувеем, встречающиеся в этом районе и не встречающиеся на Раучуа; 4 - флора Раучуа в целом; 5 - флора Эргувеема в целом.
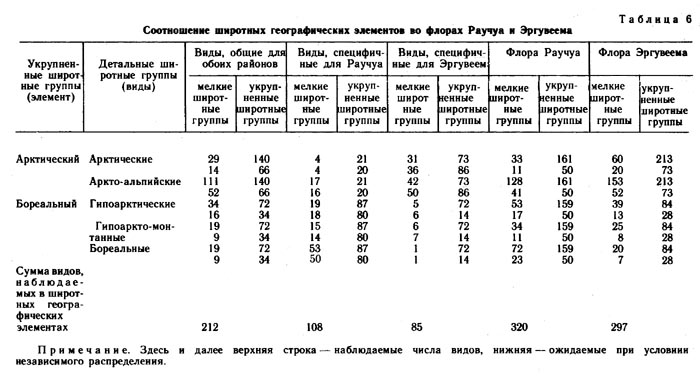
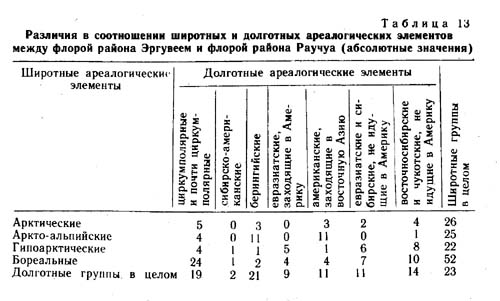
Сравнение по соотношению широтных групп (табл. 6) показывает, что во флоре Эргувеема аркто-альпийских видов 11%, а арктических на 9% больше, чем во флоре Раучуа. Иное соотношение наблюдается в группах видов гипоарктических, гипо-аркто-монтанных и бореальных. Так, во флоре Эргувеема бореальных видов на 16%, гипоарктических на 4% и гипоаркто-монтанных на 3% меньше, чем во флоре Раучуа. Отражаемые этими цифрами флористические различия двух районов в известной мере объясняются различиями в их климате с поправкой на различия в литологии, геоморфологии и ходе исторического развития природы этих районов.
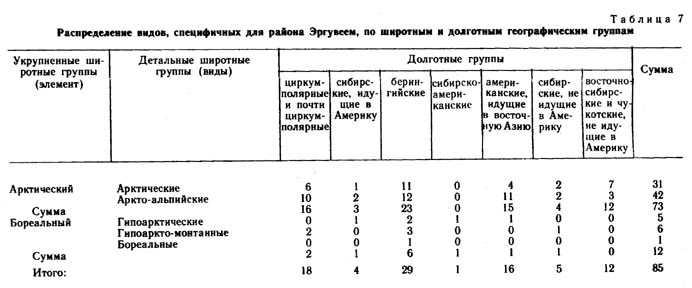
Среди видов, специфичных для района Эргувеем (табл. 7), основную массу составляют арктические и аркто-альпийские, в то время как среди видов, специфичных для района Раучуа (табл. 8), больше бореальных и гипоарктических. Вполне возможно, что в эпохи оледенений многие из бореальных видов, произрастающих в настоящее время на Раучуа, отступали на юг, но мигрировали вновь в результате потепления климата и перестройки растительного покрова.
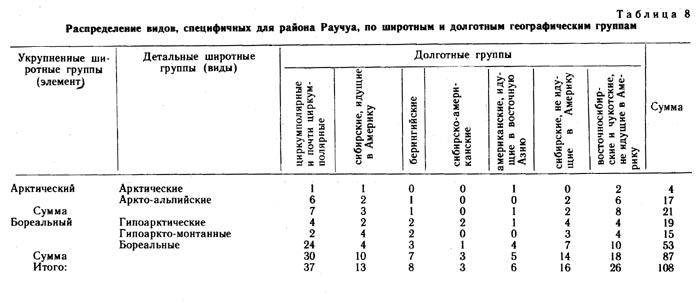
Преобладающий состав групп видов, специфичных для Раучуа, по мере сужения долготной протяженности ареала, меняется от бореального (циркумполярные и евразиатские) через бореально-гипоарктический (восточносибирские и собственно восточносибирские) до гипоарктического (чукотские). Возможно, экспансия аркто-альпийских и арктических видов на Западную Чукотку имела место во время последнего оледенения, во время же голоценового термического максимума произошло обогащение флоры бореальными видами. Какая-то часть видов арктического склада в эту эпоху могла во флоре выпасть.
В районе же Эргувеем голоценовый термический максимум, вероятно, не был связан с существенным обогащением флоры бореальными видами. Для видов, специфичных для района Эргувеем, характерно то, что по мере изменения их долготного распространения соотношение широтных групп меняется мало. На всем протяжении долготной амплитуды преобладают аркто-альпийские и арктические виды. На Эргувееме довольно много видов американских, заходящих в Азию только на Восточную и Центральную Чукотку, а западные границы ареалов таких видов, как Puccinella wrightii, Saussurea angustifolia, Solidago compacta, Aconitum delphinifolium ssp. paradoxum, проходят в непосредственной близости от района Эргувеем. Большинство американских видов, заходящих в восточную Сибирь, являются видами аркто-альпийскими и арктическими (15), и только один вид гипоаркто-монтанный. Среди видов чукотских и восточно-чукотских, специфичных для района Эргувеем, арктических видов больше (7), чем аркто-альпийских (5).
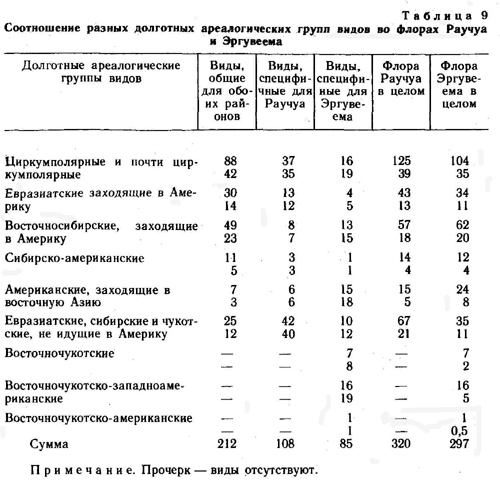 |
По соотношению долготных ареалогических групп видов выделенные совокупности отличаются также весьма существенно (табл. 9). Наибольший процент циркумполярных видов имеет место среди видов, общих для обоих районов (42%). В целом по флоре Раучуа процент евразиатско- и сибирско-западно-американских видов наибольший также в совокупности видов, общих для обоих районов, и во флоре Раучуа в целом. Виды узко распространенные, заходящие в Западную Америку, преобладают также в совокупности общих видов, но в целом во флорах процент их больше на Эргувееме. Большая часть видов американских, идущих в восточную Азию, относится к группе видов, специфичных для района Эргувеем, и в целом по флоре преобладает также на Эргувееме. В совокупности общих видов этот тип ареала представлен очень слабо.
Наибольший процент видов, не идущих в Америку, имеется в совокупности видов, специфичных для района Раучуа, в целом по флорам их больше также на Раучуа. Во флоре Эргувеема в целом их процент близок к проценту во фракции общих видов. Интересно, что процент видов восточночукотских и восточночукотско-западноамериканских в совокупности видов, специфичных для района Эргувеем, очень высок (29%), циркумполярных видов в этой совокупности 19%.
Большая часть видов, специфичных для района Эргувеем, - это виды арктические и аркто-альпийские узко распространенные, заходящие в западную Америку. Здесь нет видов гипо-арктических и бореальных с восточносибирским и чукотским долготным распространением. В совокупности видов, специфичных для Раучуа, наибольший процент приходится на виды циркумполярные (бореальные и гипоарктические), отсутствуют же виды сибирско-американские с арктическим и аркто-альпийским распространением. В совокупности видов, общих для обоих районов (табл. 10), наибольший удельный вес имеют виды с циркумполярным распространением (арктические и аркто-альпийские), наименьший - с евразиатским (бореальные и гипоарктические), а также с сибирским и сибирско-американским распространением. |

Таким образом, распределение видов разных географических групп как долготных, так и широтных по выделенным совокупностям видов (фракциям конкретных флор) весьма неравномерное. Сравнение и анализ показывают, что обе рассматриваемые флоры имеют весьма многочисленное общее ядро. Составляющие это ядро виды хорошо сформированы и являются относительно старыми таксонами, ареалы их, как правило, обширны. Различия во флорах позволяют говорить о том, что они связаны с более поздними этапами развития природы районов. На поздних этапах развития различия флор касались арктичности-бореальности и океаничности-континентальности слагающих эти флоры видов. В частности, флора Эргувеема на поздних этапах развития, вероятно, утратила ряд древнеберингийских видов континентального склада и приобрела более океанический и арктический облик, чем во времена берингийского моста суши, хотя и во времена берингийского моста она не была бореальной, а скорее, носила типичный гипоарктический облик. Ведь даже кедровый стланик не смог проникнуть по этому мосту на Аляску. Отсутствие во флоре Эргувеема следов голоценового потепления, видимо, связано не только с его отсутствием на Восточной Чукотке (такое потепление, вероятно, было), а с относительной изолированностью ввиду удаленности от бореальных флор Сибири как района Эргувеем, так и всей Восточной Чукотки. Приток бореальных видов с Аляски на Эргувеем также был невозможен, ибо в этот период уже существовал Берингов пролив. Таким образом, нет веских оснований считать, что бореальные виды, проникшие на Эргувеем в плейстоцене и голоцене, исчезли здесь в недавнее время.
В районе Раучуа в эпоху голоценового потепления, по всей вероятности, росла лиственница, образовывавшая редколесья, в пойме реки встречались чозениевые рощи и высокие ивняки, заросли же ольхового стланика встречались как на шлейфах склонов, так и на склонах сопок в местах выхода на поверхность грунтовых вод. Вполне возможно, что некоторое число видов арктического склада в результате голоценового потепления во флоре Раучуа выпали или стали чрезвычайно редкими.
В настоящее время граница лиственничных редколесий проходит в 70-80 км к юго-западу от пос. Бараниха - на южных склонах Раучуанского хребта встречаются отдельные деревья и редины из даурской лиственницы. О значительном продвижении границы леса на Север в послеледниковое время писали многие исследователи (Крючков, 1987).
Арктические и аркто-альпийские виды во флору Раучуа, вероятно, проникли в эпоху зырянского похолодания в конце плейстоцена и являются здесь более древним элементом, чем виды бореальные, быть может за исключением небольшого количества криофильно-степных видов, таких, как Carex spanioсагра, С. pediformis, С. obtusata, Festuca auriculata, Poa glauса s. 1., Pulsatilla multifida и др., которые, вероятно, сохраняются здесь с середины плейстоцена, когда на территории Бе рингии могли существовать своеобразные ландшафты криофильной тундростепной растительности, не имеющей аналогов в современную эпоху (Hulten, 1968; Юрцев, 1968, 1973; и др.). С серединой плейстоцена, по всей вероятности, связано и проникновение на Раучуа видов не степных, но имеющих разрыв ареала на Восточной Чукотке и далее встречающихся на Аляске в континентальных районах.
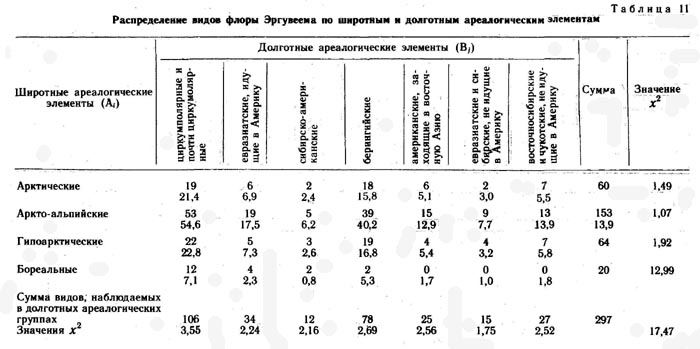
Для более полного извлечения информации о прошлом флоры и растительности исследованных районов Раучуа и Эргувеем данные по распределению видов по широтным и долготным ареалогическим элементам были подвергнуты статистической обработке методом Пирсона (табл. 11 и 12). Всего выделено 28 широтно-долготных элементов пересечения. Из таблиц видно, что распределение видов конкретных флор по элементам таблицы очень неравномерно - от 0 до 53 во флоре Эргувеема и от 0 до 49 во флоре Раучуа.
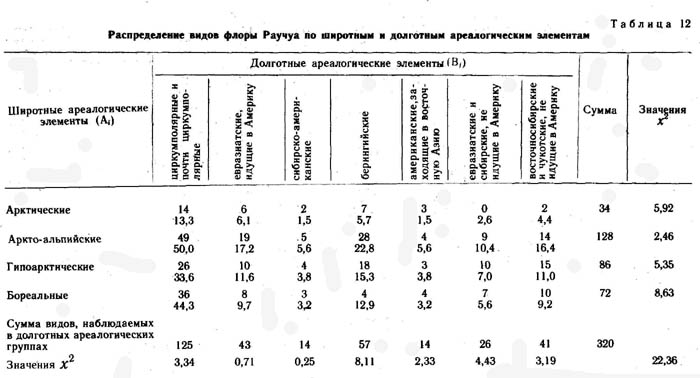
 |
В табл. 13 приведены различия в распределении видов по географическим элементам между флорой Эргувеема и Раучуа, причем значения каждого элемента табл. 13 являются абсолютной величиной разности аналогичных элементов табл. 9 и 10. Из табл. 13 видно, что наибольшее различие в группе бореальных видов в целом и в группе берингийских видов. С помощью этой таблицы выявляются широтные и долготные ареалогические. элементы флор, наиболее сильно дифференцирующие эти флоры. Такими элементами в нашем случае являются циркумполярные бореальные, берингийские аркто-альпийские, восточносибирские и чукотские бореальные. Это те географические группы видов, которые в основном и определяют различия анализируемых чукотских флор.
В табл. 11 и 12 приведены цифры, показывающие величину отклонения наблюдаемого числа видов в каждой географической долготно-широтной группе от числа видов ожидаемого в этой группе, вычисленного на основании значений краевых сумм таблицы. |
Как уже сказано выше в разделе «Методика», ожидаемые количества видов в каждом элементе пересечения определяются исходя из числа видов долготной и широтной групп (краевых сумм таблицы), к которым относится данный элемент пересечения, и общего числа видов в анализируемой флоре. Например, число наблюдаемых циркумполярных арктических видов во флоре Раучуа - 14, число всех циркумполярных видов в этой флоре - 125, всех арктических - 34, всего видов во флоре - 320. Ожидаемое число арктических циркумполярных видов должно быть (125х34)/320 = 13,3. Следовательно, отклонение от ожидаемого по этому элементу составляет + 0,7.
Во флоре Раучуа наблюдаемые положительные отклонения от ожидаемых чисел видов имеют место в группе бореальных циркумполярных ( + 8,8) и берингийских аркто-альпийских (+7,2), несколько меньше в группе восточносибирских и чукотских гипоарктических ( + 4,0) и в группе евразиатских и сибирских, не идущих в Америку гипоарктических ( + 3,0). Во флоре Эргувеема наибольшие положительные отклонения имеются в группе циркумполярных бореальных (+4,9), слабые же положительные отклонения имеются в группах берингийских арктических и берингийских гипоарктических видов. Последние, вероятно, связаны с характером видообразования в Берингии на протяжении длительного времени.
Наибольшие отрицательные значения отклонений наблюдаются во флоре Раучуа в группе берингийских бореальных видов (-8,8), циркумполярных гипоарктических (-7,6), а во флоре Эргувеема - в группе евразиатских, идущих в Америку, гипоарктических (-2,3) и берингийских бореальных (-3,3). Меньшие отрицательные отклонения во флоре Раучуа имеют цесто в группе евразиатских и сибирских, не идущих в Америку, арктических (-2,8), а во флоре Эргувеема - в группе восточносибирских и чукотских, не идущих в Америку, бореальных (-1,8) и в группе американских, заходящих в восточную Азию, бореальных (-1,7). Полученные данные совершенно однозначно показывают, что процессы видообразования и процессы миграции видов в пределах Берингии шли по арктическому и гипоарктическому типу. Растительный покров Берингии отторгал виды бореального склада, именно этим можно объяснить их дефицит во флоре Раучуа и особенно во флоре Эргувеема.
Интересно, что отклонения наблюдаемых чисел видов от ожидаемых во флоре Раучуа более существенны, чем во флоре Эргувеема. Этот факт требует специального обсуждения и мы вернемся к нему ниже по мере анализа структуры других конкретных флор.
Из полученных данных следует, что из всех арктических видов для района Раучуа наиболее «доступными» были американские, заходящие в восточную Азию, берингийские и сибирско-американские, из аркто-альпийских видов наиболее «доступными» были берингийские и евразиатские, заходящие в Америку, из гипоарктических - восточносибирские и чукотские, не идущие в Америку, а также евразиатские и сибирские, не идущие в Америку, из бореальных - циркумполярные, а также евразиатские и сибирские, не идущие в Америку. Таким образом, обмен бореальными видами, встречающимися ныне на Раучуа, между Азией и Америкой был затруднен, во всяком случае во второй половине плейстоцена. Напротив же, миграции арктических и аркто-альпийских видов, а отчасти и гипоарктических через Берингийский мост суши во второй половине плейстоцена проходили беспрепятственно.
Для района Эргувеем наиболее «доступными» были из арктических - берингийские, восточносибирские и чукотские, не идущие в Америку, а также американские, заходящие в восточную Азию, из аркто-альпийских - американские, заходящие в восточную Азию, евразиатские, заходящие в Америку, а также евразиатские и сибирские; не идущие в Америку, из гипоарктических - берингийские, а также восточносибирские и чукотские, не идущие в Америку, из бореальных - циркумполярные, евразиатские, заходящие в Америку, и сибирско-американские. Таким образом, и данные по флоре Эргувеема позволяют говорить о холодном климате Берингии по крайней мере во второй половине плейстоцена, который затруднял обмен и миграцию бореальных видов между Азией и Северной Америкой. Наиболее «недоступными» для флоры Раучуа оказались циркумполярные гипоарктические, бореальные евразиатские и бореальные берингийские, арктические сибирские и евразиатские, не идущие в Америку. Это еще раз доказывает, что экспансия арктических видов на Раучуа имела место во время существования Берингии и что климат Берингии вряд ли способствовал распространению бореальных видов и тем более возникновению здесь автохтонных бореальных видов. Во второй jполовине плейстоцена в Берингии путем видообразования возникали виды арктического, аркто-альпийского и гипбарктического склада.
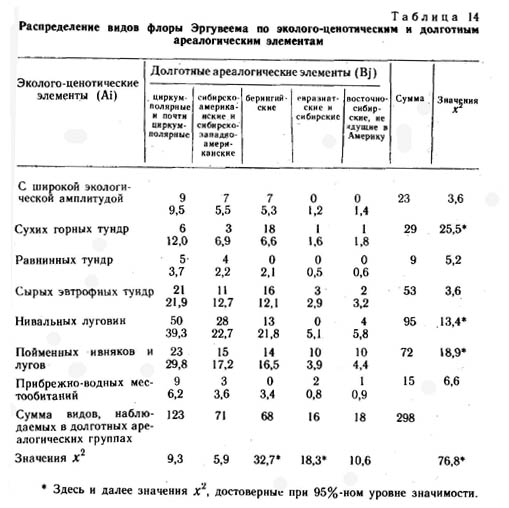 |
Наиболее «недоступными» для флоры Эргувеема оказались бореальные берингийские, евразиатские, заходящие в Америку, гипоарктические, американские, заходящие в восточную Азию, бореальные и восточносибирские и чукотские, не идущие в Америку, бореальные.
Большой интерес представляет географический анализ выделенных эколого-ценотических элементов конкретных флор, а также сопряженный анализ ареалогической и эколого-ценотической структур этих флор. Полученные данные (табл. 14, 15, 16, 17) показывают, что соотношение представителей разных ареалогических географических групп видов в разных эколого-ценотических элементах неодинаково как в пределах одной конкретной флоры, так и в сравнительном плане между аналогичными элементами разных флор. Анализ таблиц показывает, что во флоре Раучуа арктические и бореальные виды не входят в группы активных видов, имеющих в растительном покрове района очень широкую эколого-ценотическую амплитуду. Наибольший процент арктических видов в болотном комплексе, а бореальных - среди пойменных и водных видов. На Эргувееме арктические виды входят в состав комплекса видов с широкой экологической амплитудой, большой процент их также в эвтрофном тундровом комплексе и среди видов горных местообитаний. Арктические виды на Эргувееме более активны, чем на Раучуа, бореальные же виды, напротив, более активны на Раучуа. |
Процент видов, не проникающих в Америку, выше среди видов с «горной» экологией. Это, вероятно, связано с тем, что равнинные пространства осушенного шельфа для миграции горных видов представляли некоторый барьер. Кроме того, препятствием для миграции видов, обитающих в горных тундрах, являлась разница в литологии горных пород Аляски (цепи хребта Брукс в основном состоят из известняков) и Чукотки (преобладают кислые породы).
В связи с тем что арктическими и бореальными видами насыщены одни и те же эколого-ценотические комплексы, можно предположить, что растительность пойменных болот, галечников и песчаных пляжей в поймах рек, водных и эвтрофных местообитаний во флористическом отношении при климатических изменениях изменяется меньше, чем растительность склонов сопок, шлейфов без подтока минерализованных вод и равнинных увалов. Это значит, что при климатических изменениях флористический состав сухих пятнистых и щебнистых тундр изменяется в большей степени, чем флористический состав прибрежно-водной растительности, растительности галечников и песчаных аллювиальных отложений. На определенных местообитаниях в ландшафте виды растений в растительном покрове при климатических изменениях удерживаются дольше. Пойменные местообитания, например, в ландшафте работают как своеобразный накопитель и «резервуар» многих редких видов, которые могут активизироваться при соответствующих изменениях климата и существенно расширять свою эколого-ценотическую амплитуду. |
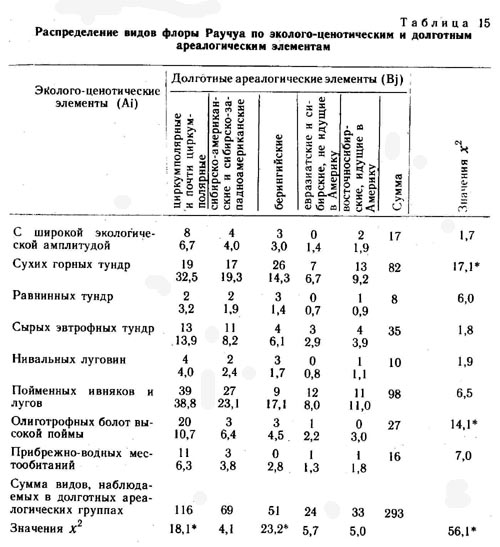
|
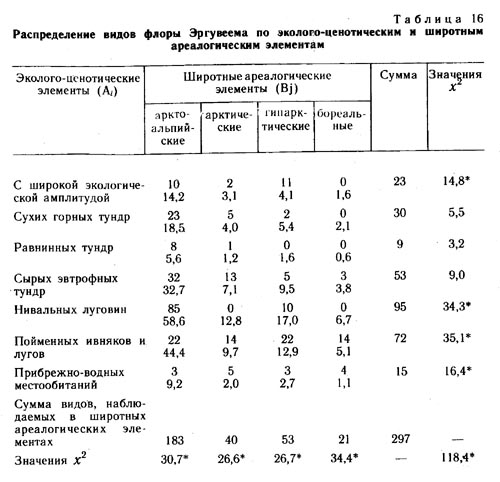 |
Интересно, что в отношении циркумполярных и почти циркумполярных видов во флоре Эргувеема наиболее богатый комплекс водных и прибрежно-водных видов (60%) и группа активных видов с горной экологией (60%), наименее богаты циркумполярными видами группа активных видов с горной экологией, а также комплекс низкоактивных видов с горной экологией и комплекс пойменных видов. Во флоре же Раучуа наибольший процент циркумполярных и почти циркумполярных видов приходится на комплексы водных и болотных видов, а наименьший - на комплекс низкоактивных видов с горной экологией.
По сравнению с долготными географическими элементами, широтные элементы по эколого-ценотическим группам и комплексам видов в пределах конкретных флор распределяются более неравномерно. Таким образом, долготная протяженность ареалов растений в большей степени связана с возрастом вида, чем широтная протяженность, так как расселение вида в меридиональном направлении меньше лимитируется экологическими факторами, чем в широтном.
Разные эколого-ценотические элементы флор имеют разную ареалогическую структуру, а следовательно, и разную историю формирования. Так, во флоре Эргувеема в комплексе видов, тяготеющих к сухим горным тундрам, несколько «завышено» число аркто-альпийских видов. Такая же закономерность прослеживается и во флоре Раучуа. В комплексе видов, тяготеющих к сырым эвтрофным тундрам района Эргувеем, число арктических видов больше ожидаемого. На Раучуа же в аналогичном комплексе «завышено» число аркто-альпийских видов. Таким образом, если на Эргувееме комплекс видов эвтрофных тундр по своей ареалогической структуре отличается от комплекса видов сухих горных тундр, то на Раучуа эти различия недостоверны. |
На Эргувееме в комплексе видов, тяготеющих к нивальным луговинам, число аркто-альпийских видов явно превышает ожидаемое.
То же самое, но в меньшей степени, наблюдается и на Раучуа. На Эргувееме в комплексе видов, тяготеющих к пойменным ивнякам и лугам, завышено представительство гипоарктических, бореальных и арктических видов, а на Раучуа в этом комплексе завышено представительство только бореальных видов. Широтная ареалогическая структура комплекса активных видов сходна на Эргувееме и на Раучуа. В обоих случаях превышение приходится на гипоарктические виды. Весьма своеобразна широтная ареалогическая структура комплекса видов болот высокой поймы Раучуа. Здесь больше ожидаемого видов бореальных и арктических одновременно.
На основании сопряженного анализа структуры конкретных флор Чукотки можно предположить, что при похолодании климата в тундровых ландшафтах бореальные виды будут дольше сохраняться в пойменных ивняках, на пойменных лугах, среди прибрежно-водной растительности и на болотах высокой поймы. В эпохи потепления часть этих видов из внутриландшафтных экологических рефугиумов может поселяться и за пределами речных долин в эвтрофных местообитаниях, на сухих участках склонов гор и равнинных увалов. При этом арктические виды сохраняют свои позиции также в составе пойменных ивняков, пойменных лугов, болот высокой поймы, в эвтрофных тундрах. При очередном похолодании арктические виды из экологических внутриландшафтных рефугиумов расселяются в сухие горные тундры. |
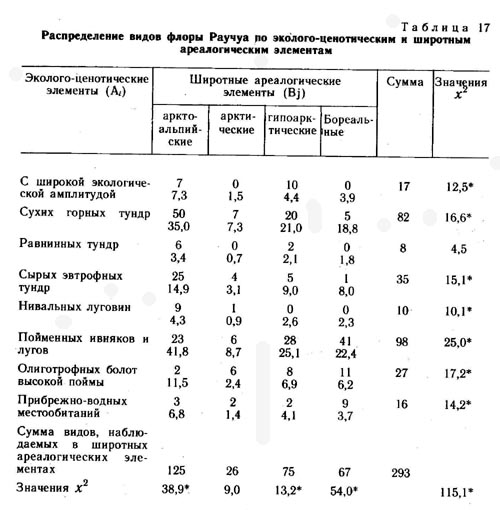
|
Анализируя сопряженность между эколого-ценотической и долготной ареалогической структурами тундровых конкретных флор, в первую очередь следует обратить внимание на виды восточносибирско-западноамериканского типа ареала - берингийские в широком смысле. На Эргувееме бёрингийские виды явно приурочены к сухим горным тундрам и избегают нивальные луговины, на Раучуа также чаще встречаются в сухих горных тундрах, однако здесь их число больше ожидаемого и в равнинных тундрах. Таким образом, в условиях берингийского сектора Арктики в неоген-четвертичное время сформировался довольно богатый комплекс видов, связанных с сухими горными тундрами. В эту группу включены и древние гольцовые элементы, распространенные в данном долготном секторе. Распределение восточносибирских видов по эколого-ценотическим элементам на Раучуа соответствует ожидаемому на основании нулевой гипотезы о независимом распределении. На Эргувееме их число завышено только в пойменных ивняках и на пойменных лугах. Примерно так же распределены в обеих флорах евразиатские и сибирские виды. Для них предпочтительнее пойменные ивняки и пойменные луга, избегают же они нивальные тундры. Эти долготные ареалогические группы стоят в обеих флорах по числу видов на последнем месте. В обеих конкретных флорах много видов, ареалы которых простираются и в западной части Северной Америки.
Проведенный сопряженный географический и эколого-ценотический анализ структуры конкретных флор Западной и Восточной Чукотки показал, что между распределением видов в ландшафте на топологическом уровне организации растительного покрова и распределением этих видов по земному шару на регионально-географическом и биосферном уровнях организации растительного покрова существуют определенные закономерные соотношения. Действительно, географические ареалогические элементы флоры по-разному представлены в эколого-ценотических элементах. Их соотношение зависит от уровня активности видов, входящих в эколого-ценотический элемент, а также от характера экологии этих видов. При климатических изменениях наиболее изменчив не набор видов определенной местности или определенного контурфитоценоза,
эколого-ценотическая активность видов. Сокращая активность при малейших климатических изменениях, виды растений пособны длительное время сохраняться в малоактивном и реликтовом состоянии на уникальных местообитаниях. Кривые распределения числа видов конкретной флоры в зависимости от их встречаемости в регулярной выборке геоботанических описаний (Галанин, 1973) наглядно показывают, сколь.велико во флоре количество видов и рас, для которых современные климатические условия далеки от оптимальных. Такие виды в ландшафте встречаются редко и весьма спорадично.
Вопрос о спорадичности распределения видов в пределах территории конкретной флоры или геоботанического района заслуживает самого пристального внимания как со стороны геоботаников-экологов, так и со стороны ботанико-географов и флористов. Понимая под спорадичностью степень нерегулярности присутствия-отсутствия видов на местообитаниях, относящихся к одному типу, мы считаем, что виды, сократившие свою эколого-ценотическую активность, будут обладать большей степенью спорадичности, чем виды, соответствующие климатической обстановке исследуемого района. Незначительной спорадичностью, вероятно, будут обладать также виды, недавно проникшие в данный район и не успевшие еще занять все подходящие для них местообитания ввиду оказываемого им ценотического сопротивления со стороны видов аборигенов. Со спорадичностью видов связана так называемая неполно-членность фитоценозов и трудности, возникающие при типизации и классификации растительности по флористическим признакам. Естественно, чем больше во флоре видов распределенных спорадично, тем более неполночленными будут фитоценозы, тем выше и неопределеннее будет их видовое разнообразие в пределах ассоциаций и групп ассоциаций.
Таким образом, для классификации растительности по флористическим признакам необходимо знание эколого-ценотического поведения видов в растительном покрове района в целом. При выделении классификационных единиц растительности по флористическому набору фитоценозов следует большую значимость придавать видам менее спорадичным, а при характеристике ассоциаций виды спорадичные отмечать особо. Мерой степени спорадичности вида в растительном покрове ландшафтного района может быть степень соответствия между его эколого-ценотическим ареалом и флористической структурой ландшафта на графовых моделях.
Флора районов среднего течения реки Цильмы, окрестностей биостанции Сыктывкарского университета
и окрестностей города Сыктывкара (сравнительный анализ)
При анализе конкретных флор этих районов были выделены широтные и долготные ареалогические элементы менее детальные, чем при анализе Чукотских флор. Это вполне оправдано тем, что для статистической достоверности в элементах таблиц сопряженного анализа должно быть значительное число видов. Использованы данные о географическом распространении видов из перечисленных в предыдущих главах источников, а также из книги «Флора Северо-Востока европейской части СССР» (1974-1977 гг.). Принципы типизации те же, что и при работе с чукотскими флорами. Сначала были выделены широтные ареалогические элементы, затем независимо от них - долготные.
Полизональные виды характеризуются ареалами, охватывающими несколько природных зон: тайги, смешанных лесов, широколиственных, лесостепи, отчасти степи и тундры. Некоторые полизональные виды заходят в зону, субтропиков, а по горным сооружениям - ив тропические регионы. К этому элементу нами отнесены следующие виды: Botrychium lunaria, Equisetum arvense, Potamogeton natans, Alisma plantago-aquatica, Agropyron repens, Phalaroides arundinaceae, Hierochloe odorata, Festuca ovina, Poa annua, Eriophorum polistachyon, Lemna trisulca, L. minor, Juncus bufonius, Urtica dioica, Comarum palustre, Potentilla norvegica, Vicia cracca, Viola arvensis, Hippuris vulgaris, Plantago major и др.
Группа аркто-альпийских видов объединена с группой арктических ввиду их малочисленности и нетипичности для изучаемых районов - все они лежат в зоне тайги. Примерами таких видов являются Phleum alpinum, Festuca rubra ssp. arctica, Omalotheca norvegica, Rumex graminifolius и др.
Гипоарктические виды: Betula nana, Deschampsia caespitosa ssp. orientalis, Pedicularis labradorica, Salix arbusculoides, Rubus arcticus и др.
Неморально-бореальные виды распространены сразу в двух зонах - в зоне тайги и смешанных лесов, а также в зоне широколиственных лесов. В этот элемент включены Matteucia strutiqpteris, Dryopteris austriaca, D. carthusiana, Equisetum sylvaticum, Abies sibirica, Pinus sylvestris, Sagittaria sagittifolia, Hydrocharis morsus-ranae, Stratiotes aloides, Milium effusum, Phleum pratense, Dactylis glomerata, Melica nutans, Carex vaginata, Paris quadrifolia, Platanthera bifolia, Dactylorhiza fuchsii, D. maculata, Geum rivale, Alchemilla monticola, Circaea alpina и др. При анализе флоры окрестностей г. Сыктывкара из неморально-бореальных видов была выделена группа неморальных видов, которые эколого-ценотический оптимум имеют в широколиственных лесах, а в зоне тайги встречаются в ее южной части, а если и проникают далеко на север, то только по уникальным местообитаниям, являющимся здесь эколого-ценотическими рефугиумами для данных видов. Такими преимущественно неморальными видами является Milium effusum, Daphne mezereum, Paris quadrifolia, лгсаеа alpina, Chaerophyllum bulbosum, Aegopodium podagraria и др.
Бореальные виды: Elymus fibrosus, Festuca beckeri ssp. sabulosa, Carex globularis, C. chordorhiza, Luzula multiflora ssp. sibirica, Salix phylicifolia, S. jenisseensis, S, bebbiana, Urtica sondenii, Polygonum viviparum, Atragene sibirica, Ranunculus lapponicus, Drosera rotundifolia, Ribes rubrum, Rubus chamaemorus, R. humulifolius, Alchemilla conglobata, A. sar-matica, Empetrum nigrum, Ledum palustre, Euphrasia vernalis, Lonicera pallasii, Cirsium heterophyllum и др.
Лесостепные или лугостепные виды в основном распространены в лесостепной и степной зонах, там они имеют эколого-ценотический оптимум, но по сухим местообитаниям с травянистой растительностью, часто по обочинам шоссе и железных дорог, заходят далеко в глубь таежной зоны. В качестве таких видов мы рассматриваем Bromus mollis, Allium angulosum, Euphorbia latifolia, Buglossoides arvensis, Echium vulgare, Veronica spuria, Filago arvensis, Cirsium setosum и др. По меридиональной протяженности ареала для конкретных флор Коми АССР мы рассматриваем семь элементов.
Циркумполярные и почти циркумполярные виды Equisetum pratense, Eq. arvense, Eq. palustre, Juniperus communis, Sparganium minimum, Agropyron repens, Deschampsia caespitosa, Milium effusum, Carex limosa, C. brunnescens, C. cinerea, Lemna trisulca, Juncus bufonius, Salix bebbiana, Rumex graminifolius, Polygonum aviculare, P. viviparum, Stellaria crassifolia, Nymphaea tetragona, Caltha palustris, Ranunculus gmelinii, R. lapponicus, Cardamine pratensis и др.
Евразиатские и европейско-западноазиатские на восток Евразии распространены до Прибайкалья. Ареалы этих видов охватывают всю или почти всю Европу и большую часть северной Азии или Сибири. К этому элементу отнесены Botrychium lunaria, В. multifidum, Pinus sylvestris, Potamogeton trichoides, Sagittaria sagittifolia, Hydrocharis morsus-ranae, Elymus fibrosus, Calamagrostis canescens, Agrostis tenuis, Carex acuta, C. leporina, C. elongata, Juncus compressus, Veratrum lobeilianum, Paris quadrifolia, Betula pubescens, Dianthus superbus, Vicia cracca и др.
Азиатские (сибирские) и азиатско-восточноевропейские виды широко распространены в северной Азии, и в Европу могут проникать вплоть до Русской равнины и Валдайской возвышенности. В этом элементе рассматриваются Abies sibirica, Picea obovata, Luzula multiflora ssp. sibirica, Salix jenisseensis, Aconitum septentrionale, Atragene sibirica, Spiraea media, Rubus humulifolius, R. sachalinensis, Cirsium setosum и др.
Европейские виды имеют ареалы, не выходящие за пределы Европы: Stratiotes aloides, Bromus mollis, Festuca beckeri ssp. sabulosa, Luzula multiflora, ssp. frigida, Allium angulosum, Salix acutifolia, S. pentandra, S. myrsinitolia, Betula nana, B. pendula, Alnus incana, Urtica sondenii, Stellaria holostea, S. nemorum, Silene tatarica, Trollius europaeus, Ranunculus auricumus, Sorbus aucuparia, Alchemilla monticola, A. sarma-tica, .Trifolium medium, T. spadiceum, Chaerophyllum bulbosum, Veronica opaca, Euphrasia vernalis, Viburnum opulus, Knautia arvensis, Campanula cervicaria и др. К этой группе примыкают виды с американо-европейскими ареалами, так как относительно флоры Коми АССР они выступают как типичные европейские виды, мигрируя с запада и юго-запада. Примерами американо-европейского типа ареала могут служить ареалы Carex nigra и Scleranthus annus.
Эндемики Северо-Востока европейской части СССР: Роа tanfiljevii, Alchemilla hyperborea, A. uralensis, A. cunctatrix, Geranium sylvaticum var. albiflorum, Euphorbia borodinii и др.
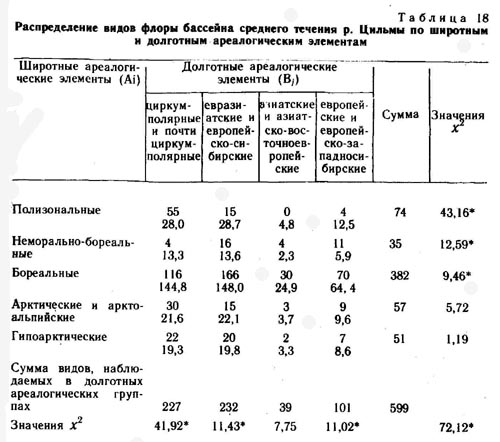 |
В табл. 18 приведены результаты сопряженного анализа широтной и долготной ареалогических структур флоры района среднего течения р. Цильмы. Суммарная величина критерия Пирсона х2 =72,12 говорит о достоверной сопряженности структур. Завышенное по сравнению с ожидаемым число циркумполярных полизональных видов говорит о том, что между долготной и широтной протяженностью ареала существует прямая пропорциональная зависимость. Именно высокая экологическая толерантность .видов обеспечивает возможность их быстрого распространения на значительные территории. Среди видов с европейским и европейско-западносибирским распространением полизональных видов, напротив, меньше ожидаемого.
«Дефицит» неморально-бореальных видов имеет место среди циркумполярных, а «избыток» - среди европейских видов. Это хорошо согласуется с положением о том, что сплошная циркумполярная зона широколиственных тургайских лесов существовала давно, и уже в середине плиоцена на востоке Евразии она была прервана таежными и тундровыми формациями. В плейстоцене в северном полушарии сформировалось несколько изолированных секторов с неморальной растительностью. Флористический обмен между этими секторами в плейстоцене был невозможен даже в эпохи межледниковий. На Северо-Востоке европейской части СССР неморальные и неморально-бореальные виды в последний раз проникли из европейского рефугиума в эпоху голоценового оптимума. Связи же с дальневосточным и южносибирским рефугиумами неморальных видов у здешней флоры в голоцене, да и в плейстоцене тоже не было. |
Бореальные виды по долготным ареалогическим элементам распределены более равномерно, чем представители предыдущих широтных ареалогических групп. Несколько больше ожидаемого здесь евразиатских, азиатских и европейских видов. Равномерность распределения указывает на то, что современные условия района больше всего благоприятны для видов таежного бореального склада, именно они наиболее свободно могли проникать сюда в послеледниковое время из южных и центральных районов Европы, с Урала и из Западной Сибири. Активная миграция этих видов в бассейн Цильмы началась раньше, чем неморальных видов и на протяжении голоцена имела более стабильный характер.
Аркто-альпийских и арктических видов больше ожидаемого в группе циркумполярных, а меньше ожидаемого в группе евразиатских видов. Число аркто-альпийских и арктических видов европейского склада близко к ожидаемому. Это позволяет предполагать, что в плейстоцене европейский сектор арктики флористически весьма существенно отличался от сибирского и здесь имел место самостоятельный центр видообразования. После разрушения ледникового покрова валдайского оледенения молодые европейские арктические и субарктические виды подверглись интенсивной миграции и интрогрессивной гибридизации. Многие из них при этом исчезли, но следы былого разнообразия проявляются в виде статистической тенденции.
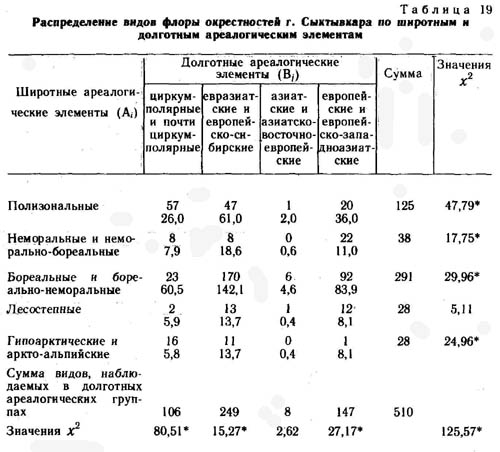 |
Гипоарктические виды по долготным ареалогическим элементам распределены равномерно. Это указывает на то, что гипоарктические виды в районе исследования находятся в эколого-ценотическом оптимуме.
По результатам ареалогического анализа флоры район бассейна р. Цильмы следует отнести к подзоне северной и крайне северной тайги гипоарктического склада восточноевропейского сектора.
В табл. 19 приведены результаты сопряженного ареалогического анализа флоры окрестностей г. Сыктывкара. Следует отметить, что здесь закономерность с распределением полизональных видов, выявленная во флоре предыдущего района, повторяется, а европейское происхождение неморальных видов видно еще более отчетливо. В группе бореальных видов существенно превышают ожидаемое число евразиатские виды и несколько меньше европейские. Это говорит о том, что в плейстоцене, и особенно в голоцене, здесь имели место два потока миграции: из Западной Сибири и с Урала и из Юго-Западной и Западной Европы. Гипоарктические и аркто-альпийские виды нa Вычегду, вероятно, проникли в плейстоцене в эпоху валдайского оледенения с Северо-Востока (с Приполярного и Полярного Урала). В голоцене многие из видов этого склада, вероятно, из флоры выпали, некоторые же сохраняются в эколого-ценотических внутриландшафтных рефугиумах. |
Результаты спорово-пыльцевого анализа показывают, что формирование флоры центральной части Коми АССР происходило на фоне неоднократных смен растительного покрова в связи с колебаниями климата. Намечается ряд фаз относительно холодного и относительно теплого климата. Холодные фазы характеризуются безлесными тундровыми ландшафтами, с теплыми же фазами связано продвижение лесов к северу, иногда значительно севернее их современного предела. Ясно, что современный растительный покров окрестностей г. Сыктывкара в основном - результат голоценовой истории, хотя часть видов могла здесь сохраняться с плейстоценового времени. Таким образом, поиски плейстоценовых реликтов здесь - очень важное направление флористических исследований. Весьма примечательно, что степень сопряженности широтной и долготной ареалогических структур флоры окрестностей г. Сыктывкара выше, чем флоры среднего течения р. Цильмы. Учитывая аналогичные данные по конкретным флорам Чукотки, можно заметить, что по мере продвижения от Арктики на юг в бореальную зону степень сопряженности широтной и долготной ареалогической структуры конкретных флор возрастает, долготное распространение видов становится столь же сильно лимитировано эколого-ценотически, как и широтное. Роль меридиональных флористических градиентов, по мере продвижения с севера на юг, становится все более соизмеримой с ролью широтных флористических градиентов.
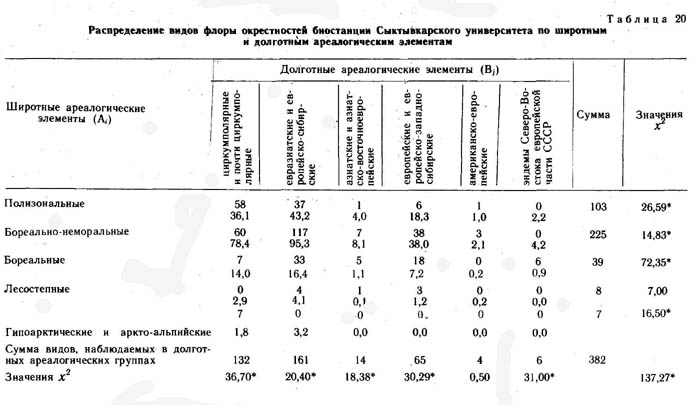
В табл. 20 приведены результаты сопряженного анализа ареалогической структуры флоры района окрестностей биостанции Сыктывкарского университета. Этот район расположен в 60-70 км восточнее г. Сыктывкара вверх по р. Вычегде. Однако, несмотря на территориальную близость, флоры различаются весьма существенно. Район биостанции имеет менее нарушенный растительный покров, здесь гораздо меньше заносных видов. Различаются и спектры экотопов этих районов. На биостанции значительную площадь занимают пойма и первая надпойменная терраса Вычегды, плакорные же местообитания практически отсутствуют, они в район исследования не вошли.
Среди полизональных видов здесь также сильно завышено число циркумполярных. Неморально-бореальные виды преобладают среди евразиатских, европейские неморальные виды во флоре в пределах ожидаемого. По всей вероятности, при формировании неморального элемента флоры окрестностей биостан-ии имели место миграции видов из южноуральского рефугиума, хотя миграции неморальных и неморально-бореальных видов с юго-запада и с запада были также довольно мощными.
Гипоарктические и аркто-альпийские виды во флоре окрестностей биостанции выпали, вероятно, в голоценовом оптимуме, осталось только семь циркумполярных видов этой группы, обладающих высокой экологической толерантностью. В целом флора окрестностей биостанции имеет неморально-бореальный характер, чем отличается от более бореальной флоры окрестностей г. Сыктывкара. Доля полизональных видов в этих флорах примерно равная. Современные климатические условия в средней части бассейна Вычегды благоприятны для видов бореального склада, особенно неморально-бореального, неморальные виды здесь представляют собой реликты эпохи голоценового оптимума. Разрушение естественного растительного покрова со сложившейся ценотической структурой приводит в первую очередь к выпадению из флоры неморальных видов. Иммигрантами же на ненарушенных местообитаниях с гораздо большей вероятностью оказываются виды полизональные. В результате антропогенного воздействия на растительный покров на месте естественных сложившихся фитоценозов возникают незамкнутые группировки растений, которые сильно обогащаются сорными заносными и рудераль-ными видами, интенсивно проникающими вдоль железных и шоссейных дорог, а также вместе с семенами культивируемых растений. В результате синантропизации на первых этапах число видов в локальных флорах увеличивается (новые виды проникли, а аборигенные еще сохраняются), но затем резко снижается за счет выпадения аборигенных видов с узкой эколого-ценотической амплитудой. В настоящее время флора бассейна Вычегды находится на первом этапе антропогенной трансформации.
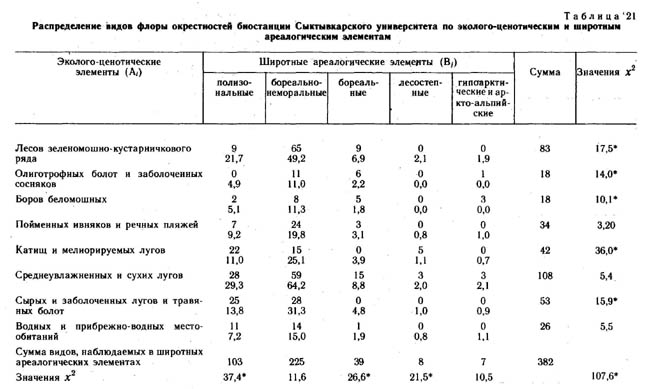
Анализ сопряженности эколого-ценотической и широтной ареалогических структур флоры окрестностей биостанции Сыктывкарского университета (табл. 21) показал, что в еловых и смешанных кустарничковых лесах число неморальных и неморально-бореальных видов достоверно превышает ожидаемое. Виды эврозональные, напротив, в этих лесах встречаются реже ожидаемого. На олиготрофных болотах и в заболоченных сосновых лесах завышено число бореальных видов и отсутствуют эвризональные. Набор видов сосняков лишайниковых (боры беломошные) по широтным ареалогическим группам сходен с таковым сфагновых болот. Распределение видов, тяготеющих к пойменным ивнякам и речным пляжам, по широтным элементам близко к ожидаемому. Эвризональные и лесостепные виды чаще ожидаемого встречаются на нарушенных лугах, в местах обработки заготовленной и вывезенной с лесосек древесины - катищах, а также на мелиорируемых лугах. Несколько неожиданно существенное отличие широтной ареалогической структуры комплексов видов сухих и среднеувлажненных лугов и комплекса видов сырых заболоченных лугов и травяных болот. Если на сухих и среднеувлажненных лугах больше ожидаемого бореальных видов, то на сырых и заболоченных лугах и в травяных болотах больше полизональных, бореальные же здесь в явном дефиците. По долготной ареалогической структуре эти типы лугов также резко отличаются друг от друга (табл. 22). На сухих и среднеувлажненных лугах больше ожидаемого европейских и европейско-западноазиатских видов при недоборе циркумполярных и почти циркумполярных. На сырых же и заболоченных лугах и травяных болотах европейских и европейско-западноазиатских видов меньше ожидаемого. Вероятно, это связано с большей стабильностью во времени комплекса видов сырых и заболоченных лугов по сравнению с комплексом видов сухих и Среднеувлажненных лугов. Полученные данные показывают, что флористический состав сырых и сухих лугов формируется по-разному, причем на сырых лугах он менее изменчив при климатических пертурбациях. Сухие и сырые луга в ландшафте средней Вычегды представляют собой совершенно разные каналы для миграции видов и сами имеют совершенно разный генезис. В связи с этим весьма интересно изучение тонкой ценотической структуры этих лугов в сравнительном плане.
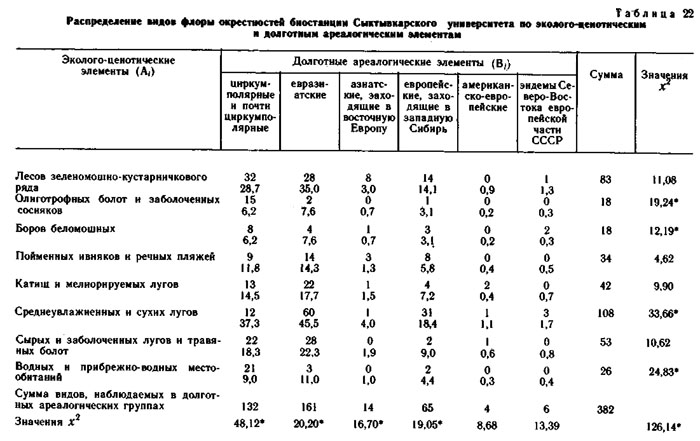
В комплексе видов водных и прибрежно-водных местообитаний явно преобладают над ожидаемым виды с циркумполярными и почти циркумполярными ареалами. Выше ожидаемого их число и в составе эколого-ценотических элементов олиготрофных болот и заболоченных сосняков. Возможно, это говорит о большей стабильности условий на экотопах с повышенной ролью широко распространенных видов. При формировании флоры еловых и смешанных зеленомошно-кустарничковых лесов сибирские флористические связи, видимо, преобладали над европейскими. Таким образом, и циркумполярные, и евра-зиатские виды на европейский Северо-Восток после оледенения проникли преимущественно из Сибири.
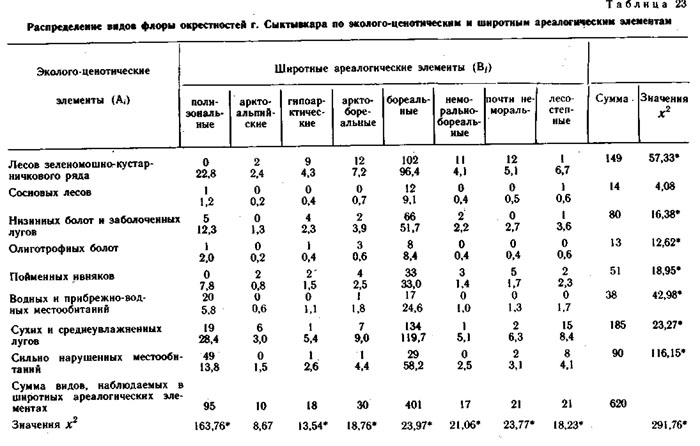
В табл. 23 дан анализ распределения видов флоры окрестностей г. Сыктывкара по эколого- ценотическим и широтным ареалогическим элементам. В комплексе видов еловых и смешанных зеленомошно-кустарничковых лесов больше ожидаемого неморальных и неморально-бореальных видов при полном отсутствии полизональных. Но, в отличие от флоры окрестностей биостанции, здесь в составе комплекса видов еловых и смешанных лесов больше ожидаемого не только сибирских, но и европейско-западносибирских видов (табл. 24). Ареалогическая структура комплекса видов сухих и среднеувлажненных лугов, заболоченных лугов, олиготрофных болот, водных и прибрежно-водных местообитаний, сильно нарушенных местообитаний в окрестностях г. Сыктывкара практически не отличается от таковой в окрестностях биостанции, что подтверждает сделанные выше выводы и предположения о характере генезиса флоры и растительности европейского Северо-Востока.
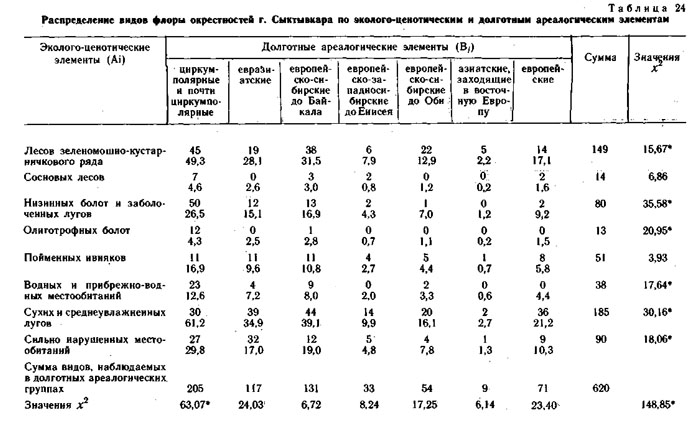
Следует заметить, что во флоре окрестностей г. Сыктывкара степень сопряженности эколого-ценотической и широтной ареалогической структур флоры выше, чем эколого-ценотической и долготной ареалогической структур. В окрестностях же биостанции это соотношение, скорее, обратное. Возможно, это связано с более дробным выделением широтных ареалогических элементов при анализе флоры окрестностей г. Сыктывкара. Вполне вероятно, что и большая антропогенная нарушенность растительного покрова в окрестностях города проявляется и в этом статистическом соотношении. Однозначно, пожалуй, одно - разные эколого-ценотические элементы конкретных флор имеют разную ареалогическую структуру, а следовательно, и разную историю формирования. Таким образом, конкретная флора в плане ее генезиса не представляет собой единую организованную изнутри систему.
Флора районов дельты реки Неман, Прегольской низменности и Вармийской конечно-моренной гряды
в Калининградской области (сравнительный анализ)
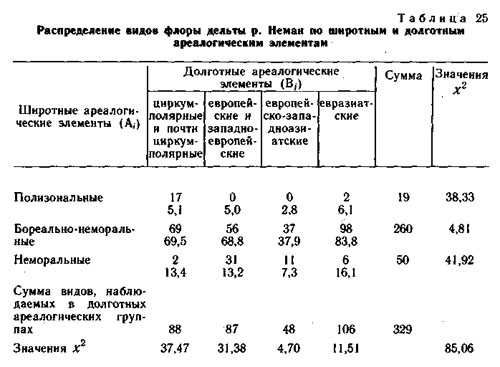 |
При анализе структуры этих конкретных флор были выделены следующие типы географических ареалов. Широкие ареалогические элементы: бореальные (широко распространены в зоне тайги и иногда заходят в соседние с ней зоны); неморальные (широко распространены в зоне широколиственных листопадных лесов и иногда заходят в соседние с ней зоны); бореально-неморальные (широко распространены как в зоне тайги, так и в зоне широколиственных лесов); аркто-бореально-неморальные, или полизональные (широко распространены в трех природных зонах); неморально-аридные (широко распространены в зоне степей и на сухих лугах или в сухих кустарниках в зоне широколиственных и подзоне хвойно-широколиственных лесов); бореально-неморально-аридные (широко распространены в трех природных зонах, по сухим лугам заходят и на север таежной зоны).
Долготные ареалогические элементы: циркумполярные и почти циркумполярные, европейские и европейские, заходящие в западную Сибирь, западноевропейские, и западноевропейско-средиземноморские, евразиатские и американско-европейские. Принципы выделения ареалогических элементов подробно изложены в предыдущих разделах монографии, там же приведены основные источники информации об ареалах видов. |
Сравнительный сопряженный анализ распределения видов по широтным и долготным ареалогическим элементам (табл. 25, 26, 27) показывает, что флора Вармийской гряды ближе по структуре к европейским флорам, чем флора Прегольской низменности и дельты Немана. Во флоре Вармийской конечно-моренной гряды несколько выше доля западноевропейских и американо-европейских видов, а также неморально-аридных и бореально-неморально-аридных. Напротив, доля циркумполярных и аркто-бореально-неморальных видов во флоре Вармийской гряды ниже. Если учесть, что конкретные флоры Калининградской области удалены друг от друга незначительно, а расстояние между флорой Вармийской гряды и флорой Прегольской низменности всего 50 км, то следует признать, что в районе р. Преголь проходит один из наиболее существенных ботанико-географических градиентов.
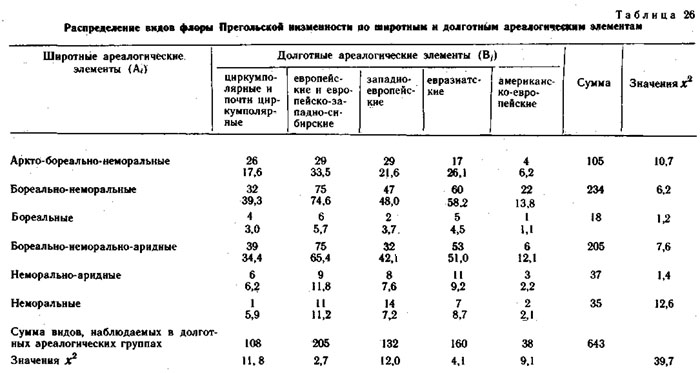
Наши данные позволяют по некоторым географическим группам ареалов видов сравнить все три конкретные флоры. Ниже приведен удельный вес разных географических групп по флорам дельты Немана, Прегольской низменности и Вармийской гряды соответственно, %:
циркумполярные: 23,2; 16,8; 11,5;
европейские: 14,7; 31,9; 31,6;
западноевропейские: 5,6; 20,5; 21,8;
евразиатские: 53,8; 24,9; 27,5;
американо-европейские: 5,3; 5,9; 7,6;
неморальные: 13,2; 18,7; 14,3;
бореальные и бореально-неморальные: 81,8; 72,5; 75,7,
Таким образом, бореальные черты флоры дельты Немана выражены значительно сильнее по сравнению с конкретными флорами, расположенными в центральной и южной частях области. Мы считаем, что для более точного установления положения и ранга ботанико-географических рубежей, проходящих через Калининградскую область и восточную Прибалтику, вообще необходимо детальное изучение ряда конкретных флор, в первую очередь это должны быть конкретная флора Самбийского полуострова, между городами Нестеров и Черняховск, на Куршской и Калининградской косе.
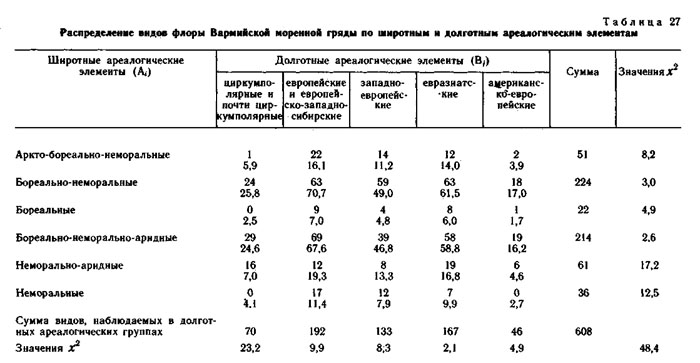
В общей сложности потрем конкретным флорам в Калининградской области нами было выявлено 850 видов и рас сосудистых растений, что составляет свыше 80% всей флоры области. Собранный гербарий хранится на кафедре ботаники Калининградского университета, часть дублетов передана в гербарий БИНа АН СССР.
Флора дельты Немана отличается бедностью, более высоким удельным весом семейства гвоздичных. Флора здесь почти в 2 раза беднее, чем в Прегольской низменности и на Вармийской гряде. Причины этого обсуждались нами в предыдущих главах. При анализе структуры этой флоры использовались укрупненные ареалогические элементы. Сопряженность между долготной и широтной ареалогическими структурами флоры здесь достоверна. Сильно завышено по сравнению с ожидаемым число полизональных циркумполярных видов. Больше ожидаемого также бореально-неморальных евразиатских, а также неморальных европейских и неморальных западноевропейских видов. Дефицит же видов имеет место в группах бореально-неморальных европейских, неморальных циркумполярных и неморальных евразиатских видов. Полученные данные показывают, что флора дельты р. Неман формировалась в голоцене миграционным путем. При этом виды приледниковой флоры, существовавшей здесь в конце плейстоцена в эпоху валдайского оледенения, очень быстро сменились видами неморального и неморально-бореального склада из западноевропейского рефугиума. Миграции растений с востока и юго-востока в район дельты Немана в голоцене практически отсутствовали.
При сопряженном анализе конкретных флор Прегольской низменности и Вармийской гряды использована более детальная или дробная система типов ареалов, причем как широтных, так и долготных (табл. 26, 27). Полученные данные показывают, что флоры по своей ареалогической структуре весьма близки друг другу. В ряде случаев различия в пределах ошибки выявления флоры. Однако для Прегольской низменности характерно значительно большее число видов аркто-бореально-неморального склада. Во флоре Вармийской гряды меньше циркумполярных и почти циркумполярных видов. Видов аридного склада чуть больше во флоре Вармийской гряды. Следует отметить, что в целом сопряженность широтной и долготной ареалогических структур флор невелика. Мы считаем, что это связано с миграционным происхождением обеих флор в недавнее голоценовое время, с одной стороны, и с сильной и давней антропогенной нарушенностью растительного покрова - с другой.
Во флоре Прегольской низменности больше ожидаемого аркто-бореально-неморальных циркумполярных видов, бореально-неморально-аридных циркумполярных, бореально-неморально-аридных евразиатских, неморальных и западноевропейских и бореально-неморальных американско-европейских. Меньше ожидаемого в этой конкретной флоре неморальных циркумполярных видов, бореально-неморально-аридных западноевропейских, аркто-бореально-неморальных европейско-сибирских и европейско-западносибирских видов. Все это хорошо согласуется с историей формирования данной флоры. Сходный характер распределения видов по элементам пересечения долготной и широтной ареалогических структур присущ и флоре Вармийской моренной гряды.
Читать следующую главу