|
Комплексные экологические исследования на стационаре "Контакт". Владивосток: Дальнаука, 1993. 281 с.
Cборник продолжает серию публикаций по стационару "Контакт" и посвящен результатам исследований, выполненных и основном на территории Колымской воднобалансовой станции (КВБС; Магаданская обл.). В него вошли статьи, представляющие результат как завершенных, так и продолжающихся исследований компонентов наземных и водных экосистем. Приводится краткий исторический очерк основных этапов становления и деятельности КВБС, рассматриваются проблемы экологических мониторинговых исследований. Представлены оригинальные материалы о структуре растительного покрова и его снизи с рельефом, результаты лихенологических, фенологических, морфологических исследований. Дается анализ фауны пауков верховьев Колымы, анализируется структура населения наземных и водных беспозвоночных, рассматриваются трофические связи мелких растительноядных млекопитающих.
Рассчитан на экологов, геоботаников, зоологов.
ISBN 5-7442-016 I -0 |
УДК 577.4 (235.33)
Ландшафтно-экологический мониторинг на стационаре "Контакт" (Верхнеколымское нагорье)
А.В. Галанин © 2011
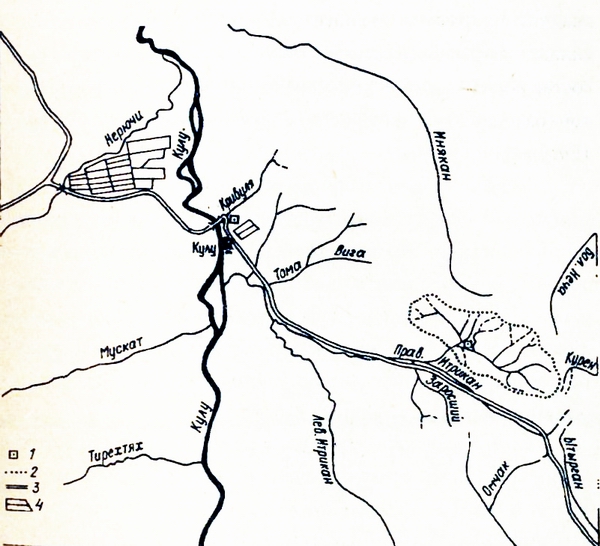
Рис. 1. Картосхема: расположение стационара Контакт в бассейне Правого Итрикана.
|
Стационар "Контакт" - ландшафтный район, весьма типичный не только для Итриканской гряды, но и для всего Всрхнеколымского нагорья (фото 1), и одно это делает его весьма перспективным для экологического мониторинга регионального ранга (рис. 1). Как известно, Верхнсколымский регион подвергается мощному антропогенному воздействию начиная с 30-х годов. Интенсивность рубок леса, лесных пожаров антропогенного происхождения и особенно роющей деятельности Человека сначала Дальстроя, а затем объединения "Северовостокзолото" в некоторые десятилетия были просто катастрофическими. К сожалению, не только в те времена, но и сегодня ни одна из комплексных программ освоения и развития больших и малых регионов страны не предусматривает систематического экологического мониторинга (Большаков и др., 1980).
Экологические последствия деятельности объединения "Севсровостокзолото" в бассейне верхней Колымы по своему размаху приближаются к последствиям освоения Байкала, а в некоторых аспектах даже явно превосходят их. Уничтожаются растительный и почвенный покровы в наиболее продуктивных долинных экосистемах, и это не может не отражаться на биосферной экологической системе всего ландшафта в целом. Немалый вред причиняет и сельское хозяйство, которое стремится уничтожить долинные комплексы там, где не содержится золото.
Уже заполнено водохранилище Колымской ГЭС, и в ближайшие гады произойдут связанные с ним климатические изменения. Попытками экологического прогноза последствий циклопического освоения природы наше общество занялось совсем недавно. Однако одной экологической экспертизы последствий осуществления тех или иных технических и сельскохозяйственных проектов мало. В процессе функционирования того или иного производства в регионе необходим систематический ландшафтно-экологичсский мониторинг - такое слежение за экологическими системами разного хорологического ранга, которое позволяло бы достоверно улавливать состояния экосистем, находящиеся за пределами их равновесия или по крайней мере за пределами допустимых флюктуационных или циклических изменений. |
Основная причина недостаточности работ по экологическому мониторингу - это неразвитость теории и методологической базы (Марчук и др., 1980; Крючков, 1987). Именно экскурс в теорию экологического и особенно ландшафтно-экологичсского мониторинга является целью настоящего очерка. Я надеюсь, что изложенные здесь положения и принципы помогут глубже понять закономерности и явления, описанные в статьях настоящего сборника. Возможно, эта статья привлечет к проблеме теории экологического мониторинга внимание молодых специалистов, которые пока все силы отдают конкретным экспериментальным исследованиям.

Фото 1. Колымское нагорье. Вид из космоса. Справа - р. Колыма, вверху р. - Кулу, в левом верхнем углу - р. Итрикан. Космоснимок из Google Earth
|
Под экологическим мониторингом вслед за Ю.А. Израэлем (1986) мы понимаем систему наблюдений, оценок и прогноза антропогенных и катастрофических естественных изменений экосистем и природной среды. Служба экологического мониторинга в перспективе должна быть постоянной и осущсствляться по программе на специальных экологических стационарах и станциях сопряженно с гидрометеорологической службой квалифицированными специалистами - экологами. Основной объект экологического мониторинга - биосфера в целом и ее пространственные подразделения - экосистемы различного ранга в соответствии с многоуровневой хорологической структурой: биосферные регионы, биосферные системы конкретных ландшафтных районов, мезокомбинации биогеоценозов и, наконец, конкретные биогеоценозы (Хекстр, 1986). Мы считаем, что наиболее перспективен экологический мониторинг на уровне биосферной экологической системы конкретного ландшафтного района, так как именно ландшафт образует основное звено в иерархии биосферных экологических систем (Демек, 1977). Дифференциация биосферной системы ландшафта на экосистемы топологического уровня (биогеоценозы) и определяют его экологическую структуру, которая может быть предметом особого ландшафтно-экологического мониторинга (фото 2, 3). Взаимодействие живой (биота) и неживой (геота) составляющих в пределах ландшафтного района, находящегося в состоянии динамического равновесия, весьма устойчиво и проявляется во внутриландшафтной дифференциации микроклимата, почвенного и растительного покровов, животного населения, биологической продуктивности локальных экосистем (биогеоценозов), химического состава атмосферы и гидросферы, генетической структуры видовых популяций, флористического и фаунистического состава конкретных биогеоценозов и т.д. В соответствии с этим могут быть созданы различные системы экологического мониторинга: климатические, почвенные, геоботанические, геозоологичсские, биопродуктивные, биогсохимические, популяционно-генетические, флористические, фаунистические и т.д. |
Проблема экологического мониторинга - это в первую очередь проблема подвижного равновесия или устойчивости природных экосистем. Согласно всеобщему принципу подвижного равновесия Ле-Шателье, всякое воздействие на равновесную систему вызывает в ней ответные изменения, которые в своей совокупности стремятся привести систему в первоначальное состояние или, по крайней мере, уменьшить изменения - погасить возмущение (Свирижев, Логофет, 1978). Благодаря отрицательным обратным связям, когда внешний импульс вызывает замкнутый круг или контур изменений, в ландшафте возникает динамическое равновесие (Демек, 1977), и устойчивость геосистемы, таким образом, есть повторяющаяся последовательность расположения элементов и блоков в пространстве и устойчивость их функционирования во времени (Дьяконов, 1981).

Фото 2. Тенькинская трасса в районе Итриканской гряды. Лиственичные леса и заросли кедрового стланика. Фото Игоря Томаса, Дмитрия Штукенберга и Олега Шахова с сайта: http://ruskiller-nsk.narod.ru/trips/kolyma-2008/kolyma-2008.htm |
Динамическое равновесие биосферной экологической системы ландшафтного района имеет место до тех пор, пока воздействие на компоненты системы не превысит некоторых пороговых нагрузок, при которых ландшафтная экосистема переходит в неустойчивое или метастабильное состояние. Таким образом, основную задачу экологического мониторинга в общем виде можно сформулировать как различение равновесного и метастабильного состояний экосистемы и определение величины пороговых нагрузок для факторов, влияющих на данную экосистему, при которых наступает "экологическая катастрофа" - система разрушается и на ее месте образуется новая экологическая система. В принципе у экосистемы любого ранга может быть несколько типов состояний:стабильное равновесие; нестабильное равновесие, при котором самый незначительный импульс может повлечь ''экологическую катастрофу"; динамическое равновесие, когда компоненты и вся система совершают сбалансированные колебания относительно постоянно развивающегося или постепенно изменяющегося некоторого инварианта состояния системы; метастабильное состояние, когда экосистема разрушается (состояние "экологической катастрофы").
Фундаментальное значение для теории экологического мониторинга имеет закон энтропии экосистем (Дьяконов, 1981), который характеризует направленность изменения системы. Например, если система противостоит разрушающему воздействию внешних факторов и сохраняет равновесие, то энтропия этой системы либо уменьшается, либо остается на прежнем уровне. Катастрофические нарушения равновесия всегда сопровождаются увеличением энергетической и структурной энтропии. Под энтропией в данном случае мы понимаем меру хаоса или общей неупорядоченности. Например, увеличение свободной энергии в экосистеме зелеными растениями - антиэнтропийный процесс. Антиэнтропийными процессами являются увеличение разнообразия экобиоморф растений и животных, увеличение видового и |
генетического разнообразия, аккумуляция энергии кристаллической решеткой глинистых минералов при их образовании из полевых шпатов и алюмосиликатов магматических пород, формирование почвенного профиля, накопление гумуса к почве, увеличение числа ярусов и биогоризонтов в экосистеме, образование дополнительных экологических ниш и т.д. Определение процессов, протекающих в экосистемах по термодинамической направленности (энтропийные или негэнтропийные), по нашему глубочайшему убеждению, и есть методологическая основа экологического мониторинга.
Экологический мониторинг невозможен, если неизвестно характерное время или хроноинтервал экосистемы. Для равновесных систем хроноинтервал - это длительность состояния нозвращения к равновесию после отклонения от него, это время релаксации системы, а для колебательных систем хроноинтервал - это полный период колебаний (Солнцев, 1981). Для биогеоценоза в ранге физико-географической фации хроноинтервал составляет время порядка 100 лет, для биосферной системы ранга ландшафтного района (элементарная биосферная система) - 1000 лет (Солнцев, 1981;Миркин, 1985), для биосферного региона ранга физико-географической области - 10 000 лет и для биосферы в целом - около 100 000 лет. Наряду с характерным временем системы при мониторинге следует оценивать также и полное время системы или хроноинтервал, равный по длительности достаточно представительному ряду характерных времен, в течение которых система успевает возникнуть, сформироваться и эволюционным путем (при возрастании негэнтропии) перейти в новое состояние - стать другой системой, более высокоорганизованной по сравнению с изначальной. Полное время характеризует систему как целостный организованный процесс. По всей вероятности, полное время экологических систем не менее чем на порядок больше их характерного времени. При экологическом мониторинге необходимо различать непериодическую и периодическую изменчивость. В настоящее время установлено, что внешними водителями ритма периодической изменчивости эко- и биосистем являются вращение Земли вокруг своей оси, вращение Земли вокруг Солнца, вращение Луны вокруг Земли, периодическое раскачивание земной оси (может быть, вокруг магнитного полюса), вращение Солнечной системы вокруг центра Галактики.

Фото 3. Ручей Правый Итрикан. Пойменный тополево-лиственичный лес. Фото Игоря Томаса, Дмитрия Штукенберга и Олега Шахова с сайта: http://ruskiller-nsk.narod.ru/trips/kolyma-2008/kolyma-2008.htm
|
Величина допустимого воздействия на экосистему различных факторов зависит от экологического резерва этой системы, так что мы можем говорить о величинах пороговых нагрузок различных факторов. В то же время состояние любой экосистемы может быть охарактеризовано как положение в системе координат, каждая из которых является частным показателем данной системы. Показателями состояния, например, могут быть количество биомассы, продуктивность, задернснность почвы, количество трофических уровней, количество экологических ниш и т.д. Для каждой экосистемы по каждому показателю имеются максимальные и минимальные допустимые значения, так что множество состояний, заключенное в интервалах значений всех показателей системы, при которых она сохраняет состояние динамического равновесия, примерно соответствует ее гомеостатическому фазовому пространству. Это и есть экологический резерв системы, от которого зависит величина пороговых нагрузок различных внешних факторов.
Биосферная система ландшафтного района представляет собой статистический ансамбль состояний, которые проходят ее территориальные компоненты и экосистемы топологического уровня за характерное время порядка первых сотен лет. В связи с этим для теории экологического мониторинга большое значение имеет эргодичесая теория физической географии (Симонов, 1972; Солнцев, 1981; Арманд, 1987), согласно которой устанавливается связь пространственных элементов и компонентов биосферной системы ландшафта с их временными изменениями. Так, например, в сериях природных компонентов и комплексов, выделяемых в пространстве в соответствии с некоторыми потоками вещества, различаются пространственные последовательности, или ряды локальных экосистем (биогеоценозов) с закономерным убыванием или возрастанием значений некоторых параметров. При изменении того же потока вещества во времени происходит сходное изменение конкретной экосистемы, и пространственный ряд |
экосистем в ландшафте может рассматриваться в качестве аналога ряда смен экосистем во времени. При использовании эргодической теории в экологии очень важно, чтобы все экспериментальные пробные площади и точки фиксированных экологических наблюдений были приведены к единому масштабу как пространства, так и времени.
Ландшафтно-экологический мониторинг невозможен вне теории биоиндикации, согласно которой структура биоты и ее отдельных синузий в ненарушенных ландшафтах в значительной степени адекватна их экологической структуре. При этом чем выше степень нарушенности биосферной системы ландшафта, тем в меньшей степени структура его биоты адекватна экотопологической неоднородности (Галанин, 1990, 1991, 1992). Метод биоиндикации применим как в геоботанике, так и в геозоологии. Индикаторами состояния экологической среды могут быть отдельные организмы - реперы, видовые ценопопуляции, многовидовые популяции организмов со сходной экобиоморфой (синузии), фито-, зоо- и биоценозы. Если принять концепцию экологической и биологической эволюции Р. Уиттекера ( Whittaker, 1972), что эволюция - это процесс увеличения биологического и экологического разнообразия на всех уровнях организации биосферы, то с помощью метода биоиндикации можно установить степень, характер и направление изменения экосистемы. В геоботанике, например, хорошо известно, чтофлюктуационные изменения сообщества растений происходят без существенного изменения видового состава за счет вариации обилия обитающих в нем видов. При изменениях, ведущих к замене одного сообщества другим, и особенно в катастрофических темпах, существенно меняется видовой состав, а также состав доминантов и эдификаторов. При катастрофических изменениях сообществ теряется их разнообразие: упрощаются вертикальная и горизонтальная структуры, уменьшается набор экобиоморф и синузий, хотя видовое разнообразие зачастую может увеличиваться, как бы компснсируя в некоторой степени потерю функционального экологического разнообразия. При эволюционных изменениях сооб ществ чем больше время эволюции, тем выше степень дифференциации внутриценотических экониш и тем более упорядоченным становится распределение особей, популяций, экобиоморф и сообществ по градиентам среды (Галанин, 1990, 1991). При повышении биологического и экологического разнообразия происходит удлинение исходных экологических градиентов среды. Для оценки структурного разнообразия сообществ могут быть использованы меры информационной энтропии (Свирижев, Логофет, 1978). К сожалению, в большинстве известных нам экологических работ количественные меры используются только при оценке видового разнообразия, в то время как наиболее важные аспекты структурного разнообразия количественно не измеряются. Хотя и меру разнообразия Шеннона, и нелогарифмические меры можно без всяких адаптации использовать при измерении разнообразия экобиоморф, синузий, возрастных состояний особей в ценопопуляции и т.д.
Важное место в теории экологического мониторинга занимает теория климакса, или устойчивого сообщества, которое находится в состоянии равновесия с условиями абиогенной среды (Миркин, 1985). В своем историческом развитии концепция климаксовой растительности прошла три стадии: климатического моноклимакса, поликлимакса и климакса мозаики. Концепция климакса мозаики находится в соответствии с другой важной экологической концепцией - теорией растительного континуума (Александрова, 1969; Трасс, 1976; Василсвич, 1983; Миркин, 1985; Галанин, 1991). Именно климаксовая растительность находится в относительном биогеохимическом равновесии с внешней средой (Галанин, 1990). При мониторинге регистрируемыми признаками климаксовой растительности являются повторяемость и устойчивость комбинаций видов в сообществах одного и того же типа климакса (Миркин, 1985); большое разнообразие экобиоморф и ценоэлементов (Галанин, 1990); высокий общий запас биомассы (Миркин, 1985); высокая чистая биологическая продукция; хорошо выраженные максимально замкнутые циклы химических элементов (Галанин, 1990); высокие показатели биоэнергетики (Миркин, 1985); выработанность почвенного профиля и отсутствие эрозии почвы; более сложная дифференциация и более сложная упаковка экологических ниш ( Whittaker, 1972); наибольшее разнообразие внутрипопуляционной дифференциации особей видов доминантов (Дыренков, 1984; Работнов, 1983).
П ри организации экологического мониторинга наибольший интерес представляют экосистемы, расположенные на ландшафтных границах (Арманд, 1987), и именно их следует рассматривать как пространственные аналоги перехода экосистем через критические состояния. Высокая уязвимость таких систем внешними факторами весьма привлекательна для ландшафтно- экологического мониторинга. В нашем случае наиболее удобной для мониторинга следует признать верхнюю границу леса в горах - так называемый подгольцовый пояс. Многие сообщества этого пояса постоянно находятся в состоянии бифуркации и при их разрушении быстро сменяются сообществами иного склада: тундра здесь может быстро замениться лесом, а леc - тундрой. Такое состояние систем в теории катастроф Тома описывается графической моделью типа "складки" (Крючков, 1987). Экосистемы с мощными эдификаторными свойствами биоценозов способны длительное время накапливать скрытые от глаза количественные изменсния, которые в определенный момент внезапно включают механизмы кардинальной перестройки. При этом чем выше эдификаторные свойства биоценоза, тем более резким и внезапным может быть его переход в новое состояние, тем глубже будет перестройка. Внезапность и глубина экологических катастроф на Севере пока что очень редко учитывается при экологических экспертизах всевозможных циклопических проектов. Аналогии с более южными районами при экспертизе северных проектов чреваты огромными потерями.
Основой ландшафтно-экологичсского мониторинга служит внутриландшафтная мозаика или рисунок биотического покрова в целом и его отдельных функциональных блоков: синузий, экобиоморф, видов. Однако хорошей основой для мониторинга внутриландшафтная неоднородность биоты и ее частей может служить только в том случае, если есть простые и точные методы ее выявления, описания, моделирования и сравнительного анализа.
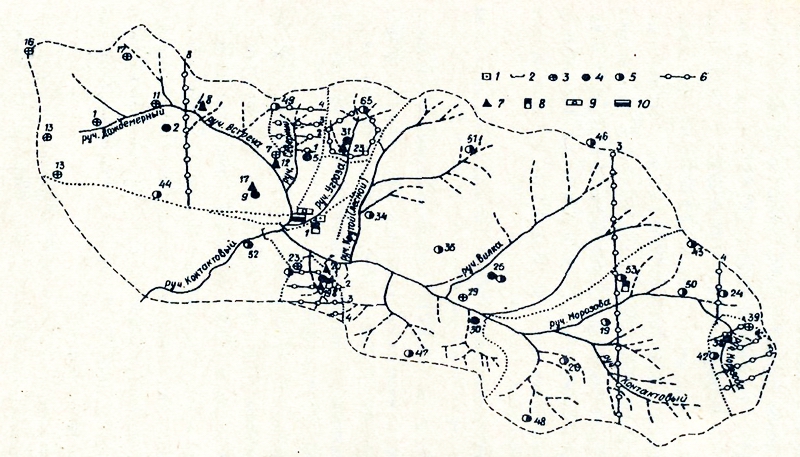
Рис. 2. Схема размещения пунктов наблюдений в бассейне ручья Стоковый Колымской водно-балансовой станции. 1 - метоплощадка, 2 - гидроствор, 3 - осадкомер, 4 - осадкомер и плювиограф, 5 - наземный дождемер, 6 - маршрутные снегомерные линии, 7 - пункт наблюдений за промерзанием и оттаиванием почвогрунтов, 9 - водноиспарительная площадка, 10 - водонепроницаемая площадка.
|
Такая методика разработана нами для анализа ландшафтно-экологической структуры растительного покрова районов конкретных флор (Галанин, 1977, 1980) и затем апробирована сотрудниками лаборатории эколого-биосферных проблем ИБПС ДВО РАН при анализе структуры животного населения (Беликович, 1988; Метод графов в экологии, 1989; Бухкало, см. наст. сб.).
Идеология нового подхода к анализу внутриландшафтной экологической структуры биотического покрова положена в основу комплексной программы многолетнего изучения ландшафтно-экологической организации биосферных систем Севера. Важной частью этой программы являются исследования на стационаре Контакт", район которого весьма типичен для зерхнеколымского нагорья. Стационар легкодоступен, на его территории много лет проводятся гидрологические и метеорологические наблюдения (Сушанский, см. наст, сб.) (рис. 2), выявлена флора сосудистых растений (Королев, Хохряков, 1988), заложена система постоянных геоботанических площадей (рис. 3), репрезентативная относительно ландшафтно-экологичсского разнообразия растительного покрова этого района (Антропова, Докучаева, 1989). Настоящий сборник является первым итогом исследований по комплексной программе, хотя и не включает всех полученных результатов - часть материалов находится в обработке. |
Основа всех экологических исследований на стационаре - система постоянных пробных площадей размером 50x50 м, заложенных в разных высотных поясах, на разных элементах мезорельефа, в разных типах растительного покрова. В своей совокупности система постоянных пробных площадей охватывает все основное разнообразие растительного покрова в 3-4-кратной повторности. Пробные площади отмаркированы, проведена их паспортизация, составлены полные списки видов растений, выполнено детальное картирование растительного покрова каждой площади, составлены геоботаничсские описания всех фитоценозов на каждой пробной площади. Ландшафтно-экологичсский мониторинг предполагает переописание пробных площадей каждые 15-20 лет.
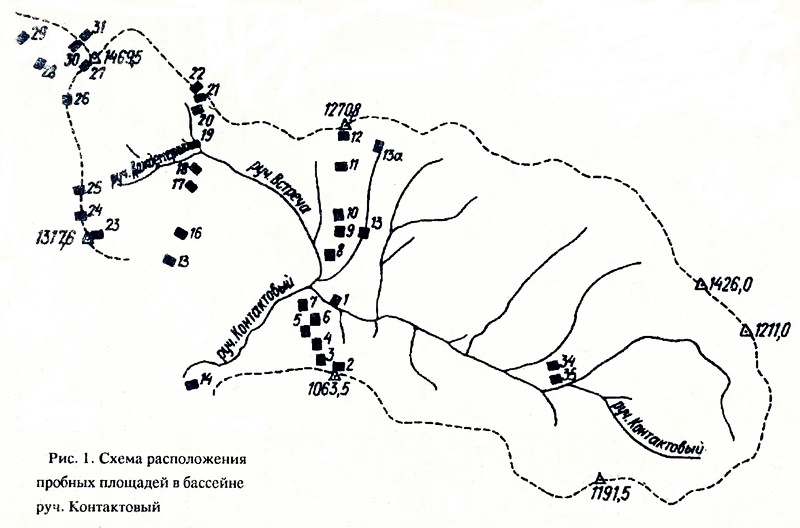
Рис. 3. Схема расположения постоянных пробных площадей на стационаре "Контакт", заложенных в 1986-1987 гг. сотрудниками лаборатории структуры растительного покрова ИБПС ДВО АН СССР. Размер площадей - 50х50 м (Антропова, Докучаева, 1989)
|
Моделями ландшафтно-экологической структуры растительного покрова, животного населения и иных биотических синузий являются ординационныс схемы постоянных пробных площадей (графы структуры), полученные путем попарного сравнения площадей на сходство-различие по характеру растительного сообщества, зооценоза или характера иных биотических синузий. В пределах района одной конкретной флоры или одной конкретной фауны сходство в наборах видов сообществ или пробных площадей всегда отражает некоторое интегральное экологическое сходство этих объектов. При этом чем больше общих видов между сообществами, тем больше и эко логическое сходство. Разумеется, зависимость экологического сходства от числа общих видов не линейная, и это было показано нами в одной из работ (Галанин, 1983). Чем шире эколого-ценотичсская амплитуда вида, тем меньше его экологическая информативность. Другими словами, если общие виды имеют узкую эколого-ценотическую амплитуду, то сходство сравниваемых сообществ больше, чем в случае, когда общие виды эвритопны и имеют широкую эколого-ценотическую амплитуду. |
За экологическую информативность вида в пределах конкретного ландшафтного района может быть принят отрицательный логарифм встречаемости этого вида в биотическом покрове данного района. Далее информативности видов суммируются при вычислении индексов сходства по Жаккару, Серснсену или иным мерам множественного сходства-различия. Однако информацию о сходстве пары сообществ несут не только те виды, которые присутствуют в обоих сообществах - это значит, что оба сообщества находятся в зоне пересечения эколого-цснотических амплитуд общих видов, но и те виды конкретной флоры и фауны, которые в данных сообществах отсутвуют - это значит, что оба сообщества лежат за пределами экогических амплитуд отсутствующих видов. Такой подход основан на признании конкретных флор и фаун целостными организованными системами, с одной стороны, и тем, что виды животных и ростений являются индикаторами среды обитания - с другой. Учитывая при расчете индексов сходства отсутствующие виды, мы также должны у читывать экологическую информативность их отсутствия (отрицательный логарифм вероятности отсутствия вида в сообществе). Такая мера использована, например, А.В. Беликович при ординации геоботанических пробных площадей ряда ландшафтных районов в бассейне Среднего Анадыря на Южной Чукотке.
Матрицы попарного сходства пробных площадей или сообщества живых организмов могут быть организованы графически, и такие графовые модели удобны для осуществления ландшафтно-экологического мониторинга, так как при повторном переописании постоянных пробных площадей мы будем получать такие же графовые модели тех же систем. Сравнение графов позволит устанавливать характер и величину изменений в интегральной ландшафтно-экологической структуре биотического покрова или его отдельных синузий. Корректные методы расчета сходства графовых структур разработаны нами ранее (Галанин и др., 1986). Кроме того, предметом мониторинга может быть степень сопряженности ландшафтно-экологической структуры различных синузий, например растительного покрова и населения почвенных беспозвоночных или почвенного покрова и населения мелких млекопитающих, и др. Наряду с этим графы можно использовать как основу для изображения и анализа ландшафтно-экологических ареалов конкретных видов, как это сделано в некоторых наших работах (Галанин, 1977, 1979). Такие ареалы также могут быть предметом специального ландшафтно-экологического мониторинга.
Таким образом, стационар "Контакт" вступает в новую стадию своего развития, и его будущее нам представляется в рамках долговременной программы экологического мониторинга. Реальной основой такого мониторинга является система постоянных пробных площадей и метод графовых моделей ландшафтно-экологической структуры биоты в целом и се отдельных синузий.
Литература
Александрова В.Д. Классификация растительности. Л.: Наука, 1969. 275 с.
Антропова Г.Л., Докучаева В.Б. Каталог пробных площадей стационара "Контакт": Препринт. Магадан: ИБПС ДВО АН СССР, 1989. Вып. 1, 2. 121с.
Арманд А.Д. Ландшафтная граница как объект мониторинга // Проблемы экологического мониторинга и моделирование экосистем. Л.: Гидромстсоиздат, 1987. С. 36-46.
Беликович А.В. Пространственная структура населения птиц Сохондинского заповедника // Структурная организация компонентов биогеосистем (сравнительный и количественный анализ). Владивосток: ДВО АН СССР, 1988. С. 79-102.
Большаков В.Н., Мамаев С.А., Ипполитов В.В. Комплексная программа "Урал - биосфера" и некоторые проблемы охраны природы на Урале // Проблемы экологии и охраны окружающей среды. Новссибирск: Наука, 1980. С. 33-41.
Василевич В.И. Очерки теоретической фитоценологии. Л.: Наука, 1983. 248 с.
Галанин А.В. Анализ распределения растений по типам местообитаний (на примере района равнинно-горного ландшафта среднего течения р. Рау-Чуа Западной Чукотки) // Ботан журн. 1977. Т. 62, №8. С. 1089-1101.
Галанин А.В. Экотопологическая структура флоры хребта Куркуре (Восточный Алтай) // Ботан. журн. 1979. Т. 64, № 10. С-1401-1413.
Галанин А.В. Флора и растительность Усть-Чаунского биологического стационара (Западная Чукотка) // Ботан. журн. Т. 65, №9. С. 1174-1187.
Галанин А,В. Сравнение флористического состава коренной и нарушенной растительности в подзоне северной тайги Коми АСССР // Ботан. журн. 1983. Т. 68, № 2. С. 174-183.
Галанин А.В. Принципы организации растительного покрова // Вестник ДВО АН СССР. 1990. № 1.
Галанин А.В. Ценотическая организация растительного покрова. Владивосток: ДВО АН СССР, 1991. 163 с.
Галанин А.В. Флора и ландшафтно-экологическая структура растительного покрова. Владивосток: ДВО АН СССР, 1992. 272 с.
Галанин А.В., Беликович А.В., Проскурина Н.С. Практические работы по биометрии: Препринт. Магадан: ИБПС ДВО АН СССР, 1986. Вып. 1, 2. 80 с.
Демек Я. Теория систем и изучение ландшафта. М.: Прогресс, 1977. 223 с.
Дьяконов К.Н. Географические законы и их физическая сущность // Геофизика ландшафта. М.: Мысль. 1981. С. 28-40.
Дыренков С.А. Структура и динамика таежных ельников. Л.: Наука, 1984. 174с.
Израэль Ю.А. О важнейших направлениях изучения и оценки антропогенных сдвигов в состоянии экологических систем // Изучение загрязнения окружающей среды и его влияния на биосферу. Л.: Гидрометеоиздат, 1986. С. 15-20.
Королев Ю.Б., Хохряков А.П. Флора сосудистых растений стационара "Контакт" (Всрхнеколымское нагорье): Препринт. Магадан: ИБПС ДВО АН СССР, 1988. 45 с.
Крючков В.В. Север на грани тысячелетий. М.: Мысль. 1987. 270с.
Марчук Г.И., Пенснко В.В., Алоян А.Е., Протасов А.В. Применение методов математического моделирования в проблеме окружающей среды // Проблемы экологии и охраны окружающей среды. Новосибирск: Наука, 1980. С. 42-53.
Метод графов в экологии. Владивосток: ДВО АН СССР, 1989. 142с.
Миркин Б.М. Теоретические основы современной фитоценологии. М.: Наука, 1985. 137с.
Работнов Т.А. Фитоценология. М: Изд-во МГУ, 1983. .384 с.
Свирижев Ю.М., Логофет Д.О. Устойчивость биологических сообществ. М.: Наука, 1978. 351 с.
Симонов Ю.Г. Региональный геоморфологический анализ. М.: Мысль, 1972. 281с.
Солнцев В.Н. Хроноорганизация географических явлений // Геофизика ландшафта. М.: Мысль, 1981. С. 40-68.
Трасс Х.Х. Геоботаника, история и современные тенденции развития. Л.: Наука, 1976. 389 с.
Хекстр Дж. П. Влияние глобально распространенных загрязняющих веществ на климат и экосистемы // Изучение загрязнения окружающей среды и его влияния на биосферу. Л.: Гидрометеоиздат, 1986. С . 26-42.
Whittaker R.H. Evolution and measurement of species diversity // Taxon. 1972. Vol. 21, № 2/3. P. 213-251.