Самоорганизация древесных ценозов
Н. А. Василенко © 2012
4.2. Пространственная структура ненарушенных
и устойчивых древостоев
Все рассмотренные выше примеры относятся к системам, состояние которых далеко от равновесного. По разным причинам эти сообщества в разной степени нарушены, и имеют разную степень хаотичности, кроме того, некоторые находятся на разных этапах самоорганизации. Какие особенности пространственной структуры можно наблюдать в системах, близких или находящихся в состоянии устойчивости, показывающих разную степень упорядоченности социальной структуры древостоя? Рассмотрим несколько примеров таких систем.
Одним из сообществ, находящихся в состоянии, близком к динамическому равновесию, является кедрово-лиственничный лес, произрастающий на склоне западной экспозиции в Сохондинском биосферном заповеднике. В пределы постоянной пробной площади входит русло ручья, его пойма и уступы надпойменной террасы. В микрорельефе встречаются россыпи гранитогнейсовых каменных глыб (Галанин, Беликович, 2004). Очевидным остается тот факт, что очень сильное влияние на пространственную структуру сообщества здесь оказывает в первую очередь особенности микрорельефа, это можно проследить на плане размещения деревьев по пробной площади (рис. 86). Кроме неоднородного характера рельефа на пространственную структуру данного древостоя, по-видимому, также накладываются и общественные отношения между деревьями: рассматриваемый древостой разделен на три дискретных группы по толщине ствола (рис. 85). Как уже отмечалось в главе 3, при составлении общей картины распределения по ценотическим классам для ценопопуляций лиственницы и кедра, входящих в древостой, характерно несовпадение пиков и провалов, особенно в правой части частотных распределений. Какова же будет картина вероятности встречи подчиненных деревьев в пределах фитогенных полей большинства особей эдификаторов в рассматриваемом древостое, с тонкой выработанной социальной структурой, но сложным эдафическим фоном? Несмотря на наличие значительных участков, не занятых деревьями, хорошо заметных на плане и связанных с внешним фактором среды, на общем поле вероятности они не проявляются, в отличие от рассмотренных нами примеров нарушенных древостоев со сложным микрорельефом.
В пределах модельного круга также незаметны какие-либо особенные пространственные образования в виде сгущений или разрежений точек (деревьев), особенно на ближнем от центра расстоянии, а зоны, где деревья отсутствуют, имеют небольшой размер (рис. 87). Это может указывать на то, что если не учитывать социальное положение и вид, к которому принадлежат деревья, то в данном древостое у деревьев нет предпочитаемых расстояний и направлений, и деревья размещены относительно друг друга в большинстве случаев равномерно. Однако, при рассмотрении характера размещения отдельных групп деревьев картина приобретает более неоднородный характер вероятности встречи особей внутри выделенных ценотических групп деревьев. Так, например, внутри группы угнетенных, с диаметром ствола 2,1-14см, деревья могут расти очень близко в направлении юго-восток – северо-запад (рис. 88), а в направлении северо-восток – юго-запад на расстоянии 1м от центра круга находится небольшой участок, где встреча особи, относящейся к классу угнетенных, маловероятна. Затем эта зона сменяется сгущением точек, вытянутых в том же направлении, что и пустоты. Вообще для поля вероятности встречи особей внутри группы угнетенных можно выделить много пустых участков, вероятно, связанных с особенностью микрорельефа и с внутренними факторами (например, некоторые из этих участков могут занимать деревья из других ценотических групп).
Группа индетерминантных деревьев с диаметром ствола от 14,1- 26см так же имеет свои особенности пространственной структуры (рис. 89) и предпочитаемые расстояния и направления. Чаще всего они могут расти на более близком расстоянии по отношению друг к другу в направлении север-юг, а вот в противоположном направлении деревья этой группы можно встретить намного реже.
В пределах модельного круга также встречаются большие участки, незанятые деревьями указанного диаметра. Вероятно, кроме внешнего фактора и фактора ценотического (т.е. занятости этих участков особями из других групп), играет роль и меньшая по сравнению с другими группами численность анализируемой совокупности деревьев. А вот лидеры в данном древостое насчитывают больше деревьев, чем группа средних. В этой группе наблюдается небольшой участок вокруг большинства особей-лидеров, где вероятность встретить еще одно дерево этой группы мала (рис. 90). Можно предположить, что такой пространственный ритм в этой группе задается уже не первичной неоднородностью среды, а является результатом происходящего в этой группе процесса выработки внутренней структуры в процессе конкуренции. Вероятно, в ходе динамики (роста деревьев) расстояние между особями внутри группы лидеров будет увеличиваться.
Еще большее расстояние относительно особей лидеров выдерживают тонкие особи, причем эта зона вытянута в том же направлении, в котором повышена вероятность встретить другую особь лидера (рис. 91). Другими словами, там, где деревья лидеры чаще растут рядом друг с другом, реже можно встретить тонкие деревья. А вот относительно особей группы индетерминантных тонкие особи могут расти и на более близком расстоянии (рис. 92), чем относительно особей-лидеров. Деревья группы индетерминантных по отношению к особям лидерам могут расти на близком расстоянии (рис. 93), но небольшая численность этой группы не дает возможности проследить наиболее предпочитаемые расстояния и направления для особей этой группы.
Обобщая полученные результаты анализа вероятностей встреч деревьев разных социальных групп по отношению друг к другу, можно предположить, что на фоне влияния сложного рельефа деревья разных ценотических групп размещены относительно друг друга неслучайно. Вероятно, наряду с выработанной внутренней упорядоченностью функциональных частей древостоя, здесь также поддерживается определенный пространственный ритм, который, по-видимому, заключается в выдержанном расстоянии и направлении между подчиненными особями и деревьями-лидерами.
Рассматривая характер размещения деревьев разных ценотических групп, можно наблюдать разное количество особей внутри классов. Встает вопрос, можно ли говорить об особенностях размещения деревьев, имея такую маленькую выборку? В этом плане мы придерживаемся мнения некоторых авторов (Эйтинген, 1949) о соизмеримой численности деревьев того или иного класса, которые произрастают в данный момент в рассматриваемой системе и характерны для нее в данных условиях и в данный момент времени.
Интересным этапом выяснения упорядоченности внутренней структуры древостоев остается вопрос изменения особенностей размещения деревьев в процессе динамики. Сохраняется ли наблюдаемый пространственный ритм, т.е. на каком расстоянии и в каком направлении от особей лидеров деревья из группы индетерминантных имеют шанс стать лидером, а деревья из группы угнетенных перейти в группу индетерминантных?

Фото 15. Кедрово-лиственничный лес (пп 5-2000) в Сохондинском биосферном заповеднике
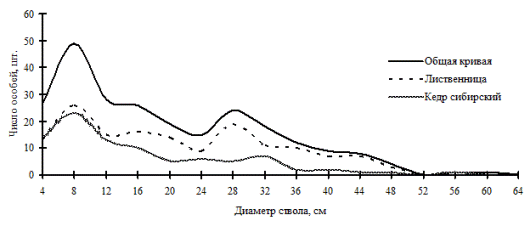
Рис. 85. Частотные распределения всего древостоя кедрово-лиственничного леса (пп 5-2000) и отдельных видов, входящих в его состав на территории Сохондинского биосферного заповедника в 2005 г. |
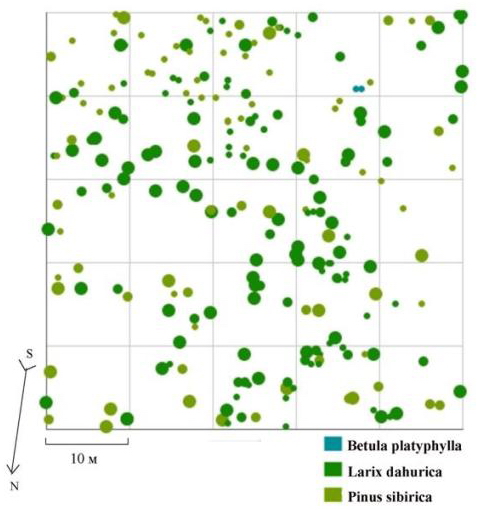
Рис. 86. Картосхема постоянной пробной площади пп 5-2000. Кедрово-лиственничный лес на территории Сохондинского биосферного заповедника |

Рис. 87. Поле вероятности встречи деревьев в пределах фитогенного поля среднестатистической особи в кедрово-лиственничном лесу (пп 5-2000) на территории Сохондинского биосферного заповедника |
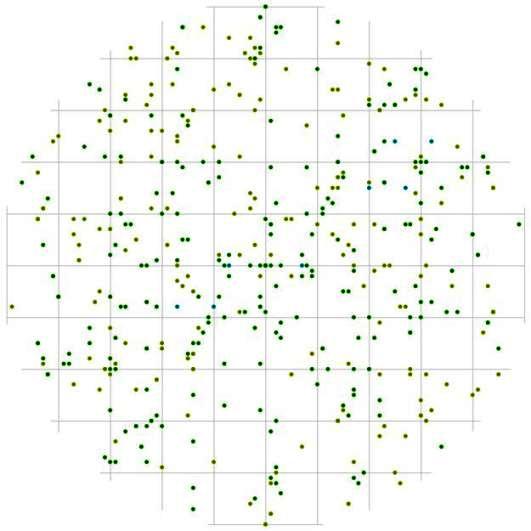
Рис. 88. Поле вероятности встречи деревьев в группе угнетенных в пределах фитогенного поля среднестатистической особи этого класса толщины в кедрово-лиственничном лесу (пп 5-2000) на территории Сохондинского биосферного заповедника |
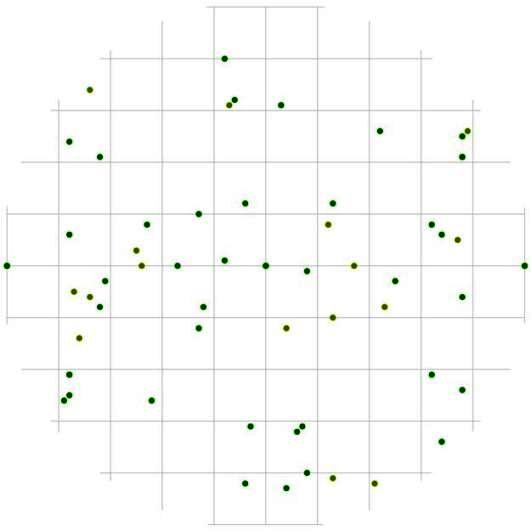
Рис. 89. Поле вероятности встречи деревьев в группе индетерминантных пределах фитогенного поля среднестатистической особи этого класса толщины в кедрово-лиственничном лесу (пп 5-2000) на территории Сохондинского биосферного заповедника |
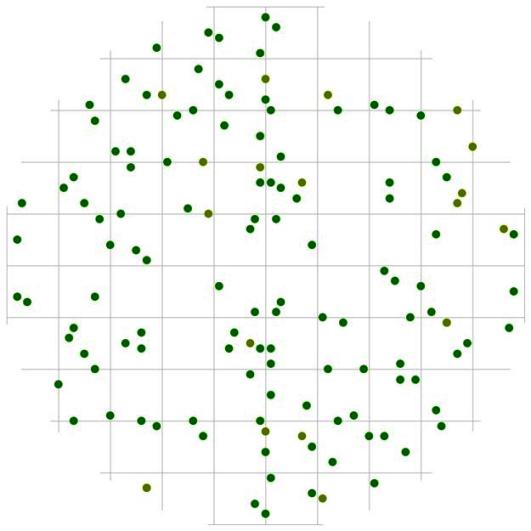
Рис. 90. Поле вероятности встречи деревьев в группе господствующих в пределах фитогенного поля среднестатистической особи этого класса толщины в кедрово-лиственничном лесу (пп 5-2000) на территории Сохондинского биосферного заповедника |
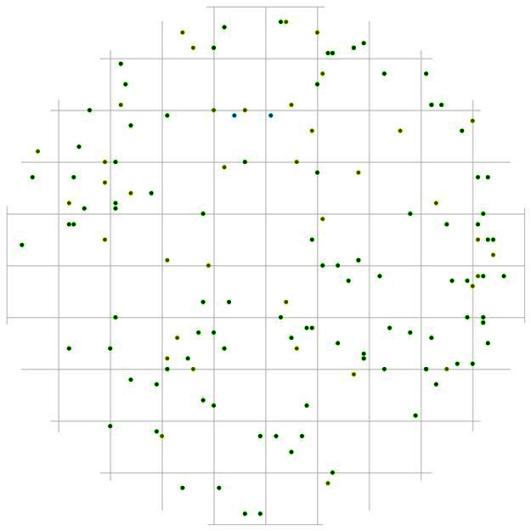
Рис. 91. Поле вероятности встречи деревьев группы угнетенных в пределах фитогенного поля среднестатистической особи господствующего класса толщины в кедрово-лиственничном лесу (пп 5-2000) на территории Сохондинского биосферного заповедника |
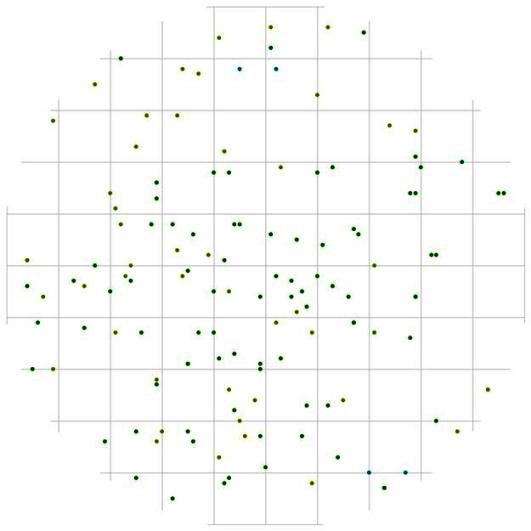
Рис. 92. Поле вероятности встречи деревьев группы угнетенных в пределах фитогенного поля среднестатистической особи индетерминантного класса толщины в кедрово-лиственничном лесу (пп 5-2000) на территории Сохондинского биосферного заповедника |
Рис. 93. Поле вероятности встречи деревьев группы индетерминантных в пределах фитогенного поля среднестатистической особи господствующего класса толщины в кедрово-лиственничном лесу (пп 5-2000) на территории Сохондинского биосферного заповедника |

Фото 16. Кедровый лес в Сохондинском заповеднике |
Другим примером растительного сообщества, находящегося в состоянии устойчивости, может служить лиственнично-кедровый лес, произрастающий в верхней части склона западной экспозиции на территории Сохондинского биосферного заповедника. Заметных рубок здесь никогда не происходило, верховых пожаров не было. Микрорельеф большей частью ровный, выходов камней на поверхность очень мало, валежа мало (Галанин, Беликович, 2004). Как и в рассмотренном выше примере, социальная структура древостоя в настоящее время имеет выработанный и устойчивый характер. Имеет место несовпадение максимумов на кривых распределения деревьев по толщине для двух ценопопуляций, составляющих данный ценоз (рис. 94), что может говорить о несовпадении во времени цикличности активизации возобновления этих видов в фитоценозе (Галанин, Беликович, 2004). Даже не проводя анализа с помощью модельного круга, из схемы размещения деревьев по пробной площади можно увидеть неравномерность в размещении особей доминирующих ценопопуляций кедра и лиственницы: особи последней встречаются в основном в левой части площади и нижних квадратах (рис. 95). Из анализа полей вероятности для всего древостоя, а также для выделенных ценотических групп деревьев видно, что этот фитоценоз выделяется из всех рассмотренных выше и тех, которые будут рассмотрены далее, тем, что во всех построенных моделях часто наблюдается размещение точек (деревьев) в углах правильной решетки, на всех расстояниях и направлениях (рис. 96-99). При этом данный факт нельзя отнести к ошибкам картирования: схема размещения строилась методом глазомерного картирования, а не измерением координат с помощью рулеток, в результате чего могло быть округление координат до целого числа.
Рассматривая построенные модели более подробно, можно выявить следующие особенности: на общем поле вероятности встречи особей в углах решетки находятся в основном крупные точки (деревья), а непосредственно в квадратах чаще всего накапливаются деревья меньшего размера (рис. 96). Очень часто незанятым деревьями остается участок до 1м в направлении восток-запад вблизи деревьев любого диаметра, в большинстве случаев это связано со средним размером каменных россыпей. Этот участок можно выделить на модельных кругах, построенных для среднестатистических особей других групп (рис. 97-99). И только для группы лидеров в этом же направлении, участок, куда не попало ни одно дерево-лидер, имеет больший размер, что уже можно, по-видимому, связать с ценотическим фактором, т.к. деревья из других классов толщины могут встречаться в пределах этой зоны (рис. 99). При раздельном рассмотрении трех групп деревьев мы видим, что особи из группы угнетенных и из группы индетерминантных деревьев, как и особи-лидеры, так же имеют вероятность находится в углах образованной решетки. На поле вероятности встречи особей из группы угнетенных относительно друг друга можно увидеть большое количество незанятых участков небольшого размера (рис. 97). Вокруг таких участков наблюдается скопление точек: это может говорить о том, что большинство тонких особей растут по краям каменистых россыпей, что также отчетливо прослеживается на картосхеме. А вот пустые участки на модельном круге для группы индетерминантных деревьев (рис. 98) можно связать, вероятно, как и в случае группы лидеров, с ценотическим фактором. Если учитывать вероятность встречи тонких особей и особей-лидеров относительно особей средней толщины, эти участки заметно сократят свою площадь.

Фото 17. Лиственнично-кедровый лес (пп 6-2000) в Сохондинском биосферном заповеднике
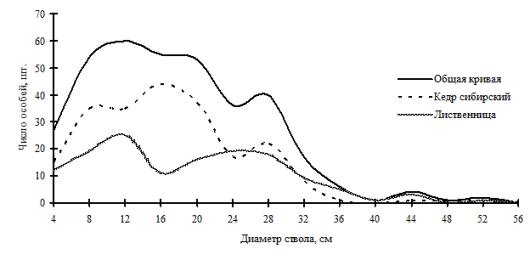
Рис. 94. Частотные распределения всего древостоя лиственнично-кедрового леса (пп 6-2000) и отдельных видов, входящих в его состав на территории Сохондинского биосферного заповедника в 2005 г. |
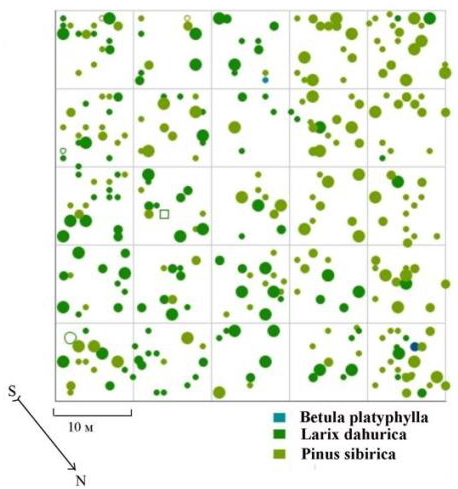
Рис. 95. Картосхема постоянной пробной площади пп 6-2000. Лиственнично-кедровый лес на территории Сохондинского биосферного заповедника |
Рис. 96. Поле вероятности встречи деревьев в пределах фитогенного поля среднестатистической особи в лиственнично-кедровом лесу (пп 6-2000) на территории Сохондинского биосферного заповедника |
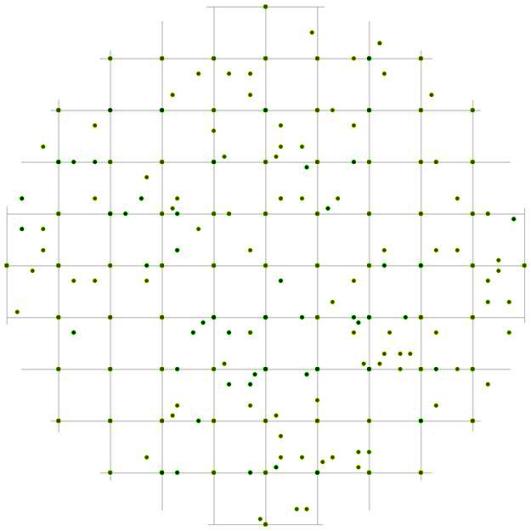
Рис. 97. Поле вероятности встречи деревьев в группе угнетенных в пределах фитогенного поля среднестатистической особи этого класса толщины в лиственнично-кедровом лесу (пп 6-2000) на территории Сохондинского биосферного заповедника |
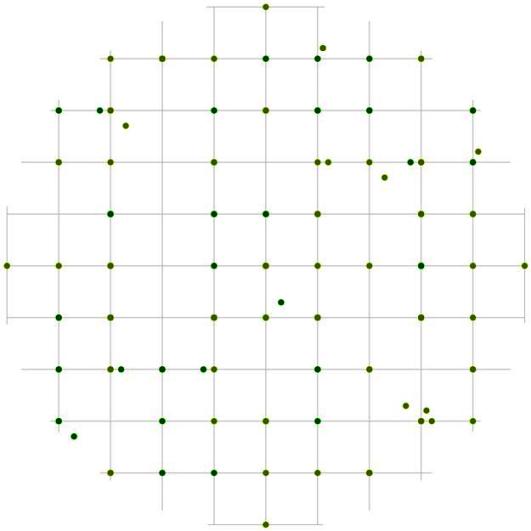
Рис. 98. Поле вероятности встречи деревьев в группе индетерминантных в пределах фитогенного поля среднестатистической особи этого класса толщины в лиственнично-кедровом лесу (пп 6-2000) на территории Сохондинского биосферного заповедника |
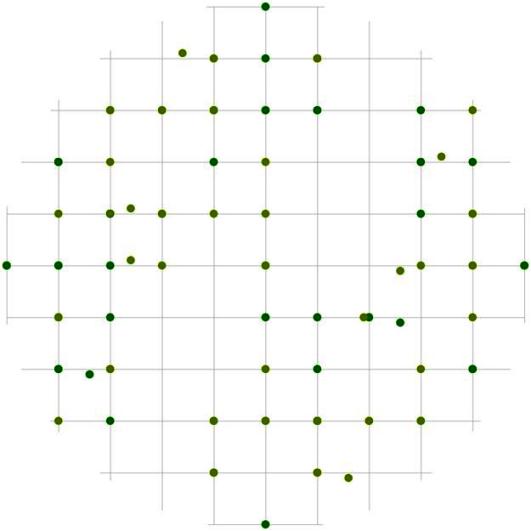
Рис. 99. Поле вероятности встречи деревьев в группе господствующих в пределах фитогенного поля среднестатистической особи этого класса толщины в лиственнично-кедровом лесу (пп 6-2000) на территории Сохондинского биосферного заповедника |
Свои особенности имеет и характер размещения деревьев разных ценотических групп. Так, например, тонкие особи относительно особей-лидеров чаще растут на расстоянии 1 м независимо от направления (рис. 100), а вот на большем расстоянии вероятность встретить особь из группы угнетенных больше в направлении северо-восток – юго-запад. Пустые участки в направлении юго-восток – северо-запад на расстоянии от 2 до 4 м, если не заняты каменными россыпями, чаще всего бывают заняты самими деревьями-лидерами. А вот на модельном круге вероятности встречи угнетенных особей относительно деревьев из группы индетерминантных очень мало пустых участков (рис. 101), практически все углы квадратов заняты точками. Интересно, что сами особи из группы со средним диаметром чаще всего могут занимать те же самые участки, что и тонкие особи, а пустые участки, не связанные с особенностями субстрата, скорее всего, будут заняты деревьями из группы лидеров. Деревья из группы лидеров могут накапливаться не только на тех же расстояниях, что и деревья из двух других групп, но и на расстояниях, на которых особи меньшего диаметра не встречаются. В размещении особей с диаметром ствола от 14,1 до 26 см относительно группы-лидеров, в отличие от предыдущих двух примеров, можно выделить много участков, не занятых точками, причем размер их в среднем равен 2м на местности (рис. 102). Пустые участки, по-видимому, в большей степени связаны с ценотическим эффектом, т.к. при учете деревьев из других ценотических групп в большинстве случаев они заняты деревьями из группы угнетенных и самими особями-лидерами.
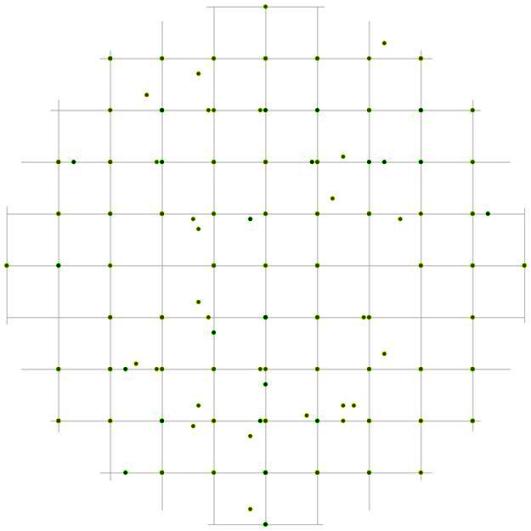
Рис. 100. Поле вероятности встречи деревьев группы угнетенных в пределах фитогенного поля среднестатистической особи господствующего класса толщины в лиственнично-кедровом лесу (пп 6-2000) на территории Сохондинского биосферного заповедника |
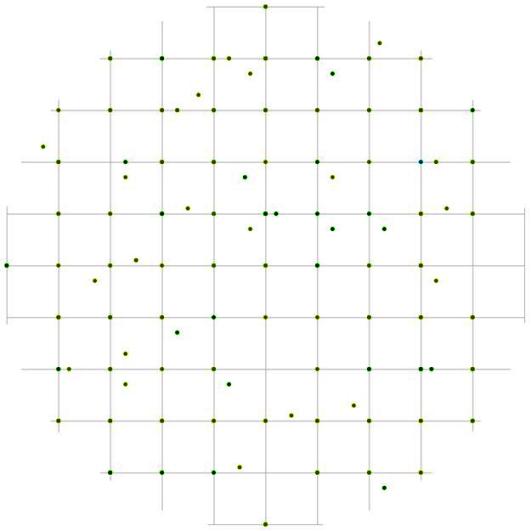
Рис. 101. Поле вероятности встречи деревьев группы угнетенных в пределах фитогенного поля среднестатистической особи индетерминантного класса толщины в лиственнично-кедровом лесу (пп 6-2000) на территории Сохондинского биосферного заповедника |
Как уже отмечалось в главе 2, сообщества, состоящие из нескольких видов деревьев, имеют сложную внутреннюю структуру, в том числе увеличено количество и состав функциональных элементов. Чем больше видов входят в состав фитоценоза, тем сложнее и разнообразней его внутренняя структура. Пространственная структура смешанных ценозов также отличается большей сложностью, чем у монодоминантных древостоев и древостоев, состоящих из двух ведущих ценопопуляций. Это также связано с неслучайным размещением особей разных видов относительно друг друга (Бузыкин и др., 1985; Галанин, 1989; Проскурина, 1991). Так, например, одни виды деревьев могут произрастать группами, другие предпочитают расти на некотором расстоянии друг от друга, но в то же время могут произрастать на близком расстоянии по отношению к другим видам.

Фото 18. Лиственично-кедровое возобновление на месте вырубки и гари в Сохондинском заповеднике |
Примером таких сложных многокомпонентных систем, как в плане наличия в составе нескольких видов, так и сложной социальной структуры древостоя и сложной пространственной структуры, может служить кедровник с елью и липой разнотравный, произрастающий на склоне юго-восточной экспозиции в заповеднике «Бастак». В древостое здесь по числу стволов преобладают несколько пород: пихта белокорая, кедр корейский, липа амурская и ель аянская. Этот древостой имеет очень сложное строение как по распределению особей по ценотическим классам, так и по характеру размещения (рис. 103-104), на особенности пространственной структуры здесь также оказывает сильное влияние характер микрорельефа, на которое накладывается взаимовлияние деревьев разных пород и разной толщины. Неоднородность в размещении особей разных пород можно проследить на схеме размещения деревьев по пробной площади (рис. 104). Если деревья одних видов (таких как пихта белокорая, кедр корейский, клен моно и др.) распределены практически по всей площади, занятой данным фитоценозом, другие (например, особи березы каменной) растут только в определенной части ценоза.
На общем модельном круге вероятности встречи особей деревьев относительно друг друга можно увидеть большое количество участков скоплений точек (деревьев), между которыми прослеживаются пустые участки, имеющие в основном небольшие размеры (рис. 105). Скопление точек вблизи центра круга может говорить о том, что часть деревьев, произрастающих в данном фитоценозе, может расти на очень близком расстоянии друг от друга. Однако это характерно не для всех видов деревьев, входящих в состав древостоя, а также не для всех групп толщины. Из каких же особенностей в размещении особей разных пород и размеров складывается общая картина распределения вероятностей? |
При раздельном рассмотрении характера размещения деревьев внутри социальной группы угнетенных особей заметно, что на близком расстоянии друг от друга чаще всего могут встречаться особи пихты белокорой и клена желтого, в направлении юго-восток – северо-запад на расстоянии до 0,5 м (рис. 106). В этом направлении можно встретить скопление точек и на больших расстояниях от центра: по-видимому, это те расстояния, которые чаще всего встречаются между самими группами. В этом древостое во всех ценотических группах активно идет процесс изреживания, что можно увидеть на рис. 43. Можно ли связать наши предположения о прохождении избирательного отпада непосредственно внутри групп с особенностями пространственного размещения живых и сухих деревьев? Наибольшее число особей отпада наблюдается в группе угнетенных деревьев, внутри этой группы вероятность встречи сухих деревьев распределена более равномерно в пределах круга. В целом, наблюдая за размещением сухих и усыхающих деревьев из группы угнетенных (сухие и усыхающие деревья обозначены на рисунках пустыми кружочками) относительно живых деревьев того же диаметра, можно заметить, что их можно встретить рядом друг с другом очень часто. Эта вероятность больше, чем вероятность встретить сухие или усыхающие тонкие деревья рядом с деревьями большего диаметра относительно этих групп. Сухие деревья из группы угнетенных на близком расстоянии встречаются чаще для особей группы индетерминантных, чем для особей-лидеров.
Такую закономерность можно проследить и на общей схеме размещения, где сухие особи из группы угнетенных чаще встречаются рядом с живыми деревьями того же размера, либо рядом с живыми деревьями группы индетерминантных особей. Это говорит о том, что избирательная форма отпада в этой группе, по-видимому, происходит как в результате взаимодействия особей внутри группы, так и в результате воздействия на них более крупных особей.
В размещении живых деревьев из группы инедетрминантных так же как и для группы тонких существует некоторая вытянутость вероятности встречи деревьев в направлении юго-восток – северо-запад (рис. 107). Однако на ближних к центру расстояниях чаще можно встретить пустые участки, а вот наибольшее скопление точек, т.е. повышенную вероятность встретить соседнюю особь того же размера, можно обнаружить на больших расстояниях от центра круга, начиная от 4 м. Большая не занятая деревьями площадь и количество пустых участков ближе к центру модельного круга могут быть связаны с внешним фактором, но в этом случае большая часть этих зон будет занята особями из других ценотических групп. Можно предположить, что здесь сказывается ценотический фактор, исключающий рост деревьев из группы индетерминантных относительно друг друга на этих расстояниях и в этих направлениях. Что касается сухих деревьев, то чаще всего сухое дерево с диаметром от 18,1 до 30 см можно встретить на расстоянии от 3 м и более, а вот по отношению к особям-лидерам сухие деревья средней толщины могут расти на меньших расстояниях (рис. 111), что может говорить о жесткой конкуренции между деревьями из двух этих групп.
В группе лидеров особи могут расти на близком друг к другу расстоянии, примерно до 1м, однако чаще всего эта вероятность складывается за счет разных пород деревьев (рис. 108), причем зона повышенной вероятности встречи подчиненной особи вытянута в направлении восток-запад. А вот в направлении север-юг вокруг большинства деревьев с диаметром больше 30см встречаются пустые участки: здесь вероятность встретить особь того же диаметра низкая, однако в этом направлении и на расстоянии от 0,5 м до 2,3-3 м часто можно встретить деревья из группы угнетенных и индетерминантных.
Интересно, что особи-лидеры чаще растут на меньшем расстоянии друг от друга, чем от деревьев из других групп. Так, особи из группы угнетенных и индетерминантных выдерживают некоторое минимальное расстояние, на котором вероятность встретить деревья этих групп вблизи особей лидеров мала (рис. 109 и рис. 111). Для группы тонких деревьев это расстояние около 0,5 м, а деревья средней толщины по отношению к лидерам чаще всего растут на расстоянии от 1 м и более. Наблюдаемые особенности пространственной структуры данного древостоя, по-видимому, могут говорить о неслучайности в размещении не только особей разных пород, но и разных социальных групп. Эта неслучайность может выражаться в особенностях пространственного размещения особей разных видов и разного ценотического статуса относительно друг друга. «Природа не терпит пустоты», этим, вероятно, и определяется порядок в живых системах: эти пустоты всегда будут заняты элементами другого порядка и положения в рассматриваемой системе.
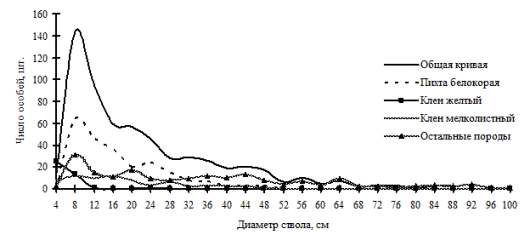
Рис. 103. Частотные распределения всего древостоя кедрового леса (01-2002) и отдельных видов, входящих в его состав на территории заповедника «Бастак» в 2002г. |
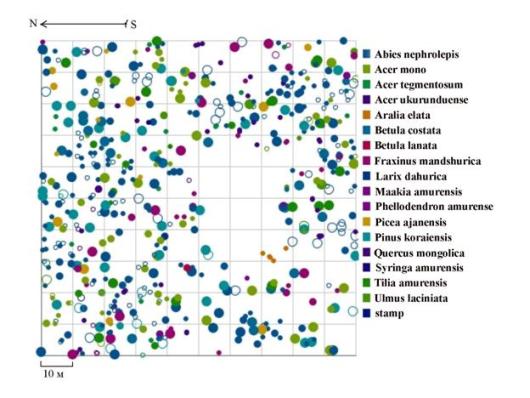
Рис. 104. Картосхема постоянной пробной площади пп 01-2002. Кедровник с липой и елью, произрастающий на территории заповедника «Бастак» |

Рис. 105. Поле вероятности встречи деревьев в пределах фитогенного поля среднестатистической особи в кедровом лесу (пп 01-2002) в заповеднике «Бастак» |
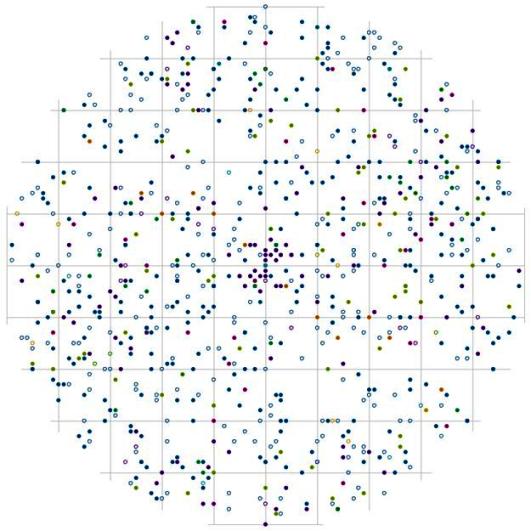
Рис. 106. Поле вероятности встречи деревьев в группе угнетенных пределах фитогенного поля среднестатистической особи этого класса толщины в кедровом лесу (пп 01-2002) в заповеднике «Бастак» |
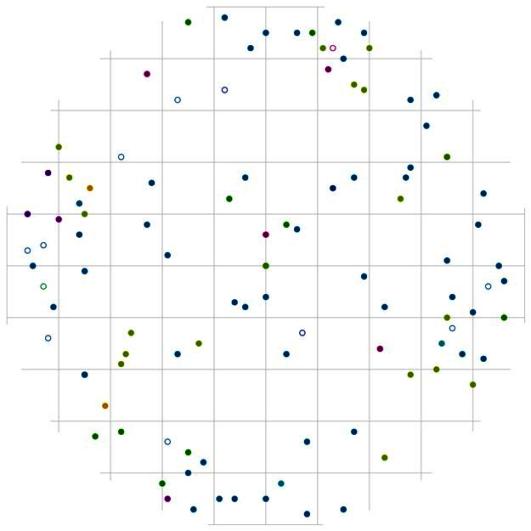
Рис. 107. Поле вероятности встречи деревьев в группе индетерминантных в пределах фитогенного поля среднестатистической особи этого класса толщины в кедровом лесу (пп 01-2002) в заповеднике «Бастак» |
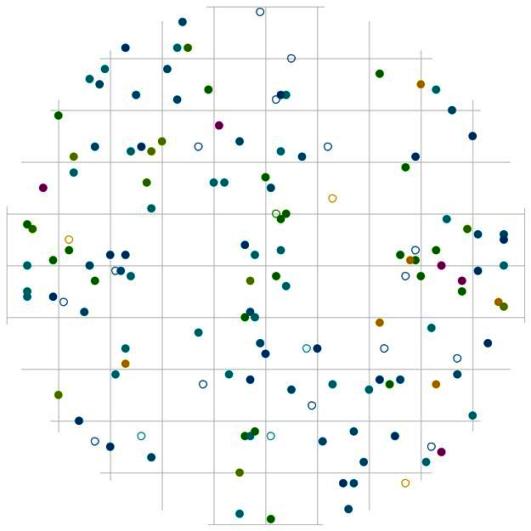
Рис. 108. Поле вероятности встречи деревьев в группе господствующих в пределах фитогенного поля среднестатистической особи этого класса толщины в кедровом лесу (пп 01-2002) в заповеднике «Бастак» |
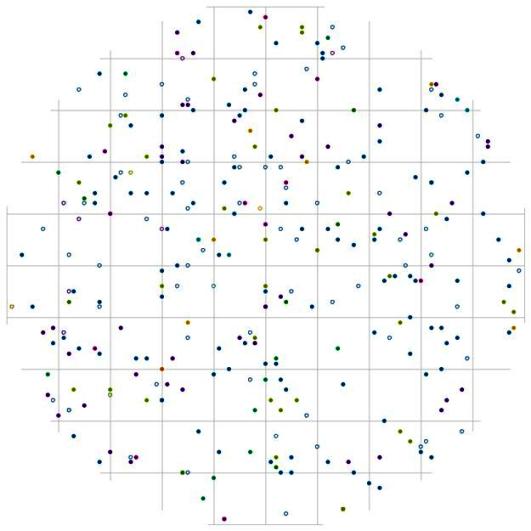
Рис. 109. Поле вероятности встречи деревьев группы угнетенных в пределах фитогенного поля среднестатистической особи господствующего класса толщины в кедровом лесу (пп 01-2002) в заповеднике «Бастак» |
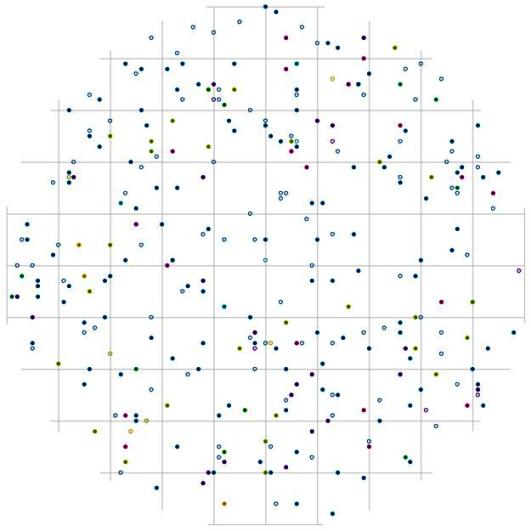
Рис. 110. Поле вероятности встречи деревьев группы угнетенных в пределах фитогенного поля среднестатистической особи индетерминантного класса толщины в кедровом лесу (пп 01-2002) в заповеднике «Бастак» |
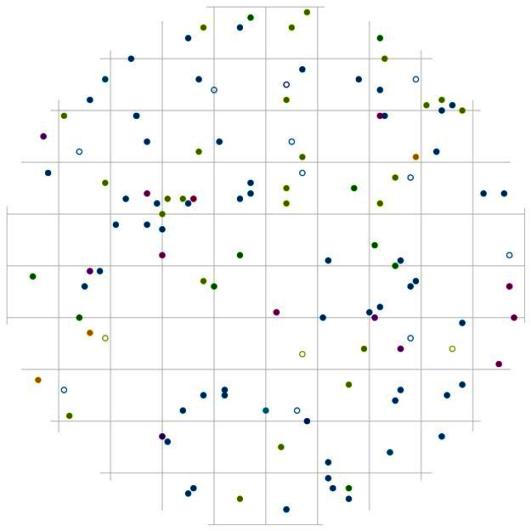
Рис. 111. Поле вероятности встречи деревьев группы индетерминантных в пределах фитогенного поля среднестатистической особи господствующего класса толщины в кедровом лесу (пп 01-2002) в заповеднике «Бастак» |

Фото 19. Н.А. Василенко в Сихотэ-Алинском заповеднике на кордоне Венера |
Еще одним примером многокомпонентной системы может служить пихтово-кедровый лес, произрастающий на склоне северо-западной экспозиции в заповеднике «Бастак». В древостое здесь по числу стволов преобладает пихта белокорая, береза ребристая и клен желтый. Как уже говорилось выше, этот пример, по-видимому, отражает также факт неслучайного характера размещения особей разных пород относительно друг друга, который является результатом взаимодействия между компонентами системы. Этот фактор накладывается на изначальное влияние среды и микрорельефа на произрастающие породы деревьев, которые могут занимать разные участки рассматриваемого фитоценоза и встречаться в той или иной части площади в большом количестве (рис.113). Так, например, как можно увидеть из схемы размещения деревьев в пределах постоянной пробной площади, особи пихты белокорой в основном размещены более равномерно, особи березы желтой чаще растут в верхней части изучаемого контурфитоценоза, а вот деревья клена желтого в основном сосредоточены в нижнем углу площади.
В рассматриваемом древостое, как и в предыдущем древесном ценозе, было выделено три ценотические группы (рис. 112). В этих двух смешанных контруфитоценозах большое число древесных пород участвует в примеси; при небольшой численности они также участвуют в образовании выделенных нами групп деревьев по толщине. Можно ли найти сходство в строении древостоев (при общей их схожести по основному морфологическому показателю) в плане пространственного размещения деревьев разных социальных групп и разных пород? Общими в данном случае могут быть одинаковые расстояния и направления, на которых предпочитают расти особи разных пород и толщины в кедровнике с елью в пихтово-кедровом лесу.
Уже на общем модельном круге можно заметить различие и сходство с предыдущим примером (рис. 114). Сразу же здесь отметим, что рассматриваемые пробные площади имеют разные размеры. Одной из общих черт является большая вероятность встречи особей разных пород и разной толщины на расстоянии 1 м, в направлении северо-восток – юго-запад от центра, а также небольшие участки в обратных направления юго-восток-северо-запад на расстоянии 0,5 м, куда не попало ни одно дерево в двух рассматриваемых ценозах. Можно сказать, что, несмотря на разницу в размере изучаемых контурфитоценозов, а также разный тип растительности, наблюдаются сходные черты в размещении особей относительно друг друга. А вот одним из различий является характер размещения и количество сухих деревьев в разных ценотических классах, что можно увидеть на рис. 43-44 и на общих модельных полях (рис. 105-114). Однако поменяются ли общие черты в размещении деревьев внутри групп и между особями разной толщины, если особенности характера размещения рассмотреть более детально?
Действительно, при раздельном рассмотрении характера размещения деревьев разных классов толщины обнаруживаются индивидуальные для данного ценоза черты. Для группы угнетенных особей большая вероятность встречи особей из этой же группы находится вблизи центра: это говорит о том, что тонкие деревья очень часто растут на малом расстоянии друг от друга. В отличие от предыдущего древостоя, тонкие особи рассматриваемого пихтово-кедрового древостоя чаще растут в направлении северо-восток – юго-запад на расстоянии от 0 до 1м от действующей особи (рис. 115). Причем такая картина вероятности задается в основном деревьями клена желтого и березы желтой, особи других видов (например, тонкие особи пихты белокорой, липы амурской и других пород) чаще выдерживают большее расстояние относительно других тонких деревьев. Как уже отмечалось выше, в изучаемом древостое наблюдается меньшее число сухих особей, однако, как и для всех рассмотренных в работе контурфитоценозов, большую часть составляет группа угнетенных деревьев.
Изучая характер размещения сухих деревьев внутри этой группы, можно отметить, что расстояния, на которых встречаются сухие особи с диаметром от 2,1 до 18 см, несколько больше, чем в кедровнике с елью. В пихтово-кедровом лесу такие особи можно встретить на расстоянии 1 м от действующих особей того же диаметра, и особенно в направлении восток-запад, а на расстоянии меньше 1 м - в направлении юго-восток и юго-запад по отношению к живым деревьям того же диаметра. Судя по характеру размещения сухих деревьев из группы угнетенных относительно других ценотических групп деревьев (рис. 118-119), можно предположить, что дискриминационная форма отпада угнетенных особей идет не только внутри группы, при взаимодействии тонких деревьев между собой, но и под влиянием более толстых деревьев.
В отличие от группы тонкомера, деревья внутри группы индетерминантных стремятся выдержать определенное минимальное расстояние (рис. 116). Эта группа немногочисленна, но также состоит из нескольких пород деревьев, которые и образуют совместно такой характер размещения. Судя по частотному распределению сухих деревьев в данном древостое (рис. 44), на смену усохшим деревьям-лидерам пришли особи из группы среднего диаметра. Это предположение можно сделать также на основе особенностей размещения сухих и живых деревьев лидеров: расстояние между ними часто бывает небольшое. По-видимому, рядом с сухими особями господствующего класса росли особи из группы индетерминантных, которые в последствии и перешли в группу лидеров (рис. 113-117). При этом, вероятно, внутри рассматриваемого древостоя образовалось несколько групп деревьев-лидеров. Как и внутри группы тонких деревьев, особи-лидеры могут расти рядом, но, как показывает рис. 117, это могут быть деревья-лидеры разных видов. На близком расстоянии от среднестатистической особи-лидера чаще всего можно встретить особь пихты белокорой.
При этом вероятность встречи соседних деревьев лидеров вытянута в направлении восток-запад, а вот в направлении север-юг от центра поля (т.е. от большинства деревьев с диаметром более 26 см) существует пустые участки протяженностью около 2 м, где вероятность встречи соседней особи-лидера минимальна. Такие же незаполненные деревьями участки можно встретить во всех направлениях, но на больших расстояниях, в среднем от 2 до 3 м. Большинство из этих участков чаще всего заняты особями других ценотических классов (рис.118 и рис. 120). Например, особи из группы угнетенных деревьев могут расти на любом расстоянии и направлении от деревьев из группы лидеров (рис. 118). Особи из группы индетерминантных чаще всего встречаются на более близком расстоянии к особям-лидерам в направлении северо-восток и северо-запад (рис. 120). Оставшиеся же пустоты можно связать с особенностями микрорельефа, а также с ценотическим фактором, исключающим произрастание подчиненных особей в пределах фитогенных полей особей из класса господствующих.
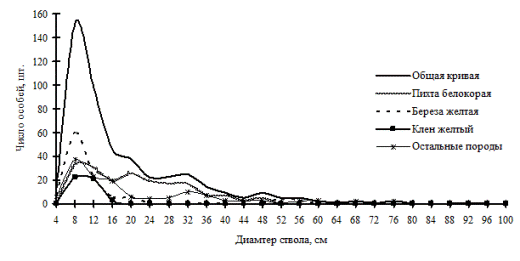
Рис. 112. Частотные распределения всего древостоя пихтово-кедрового леса (02-2002) и отдельных видов, входящих в его состав, на территории заповедника «Бастак» в 2002г. |
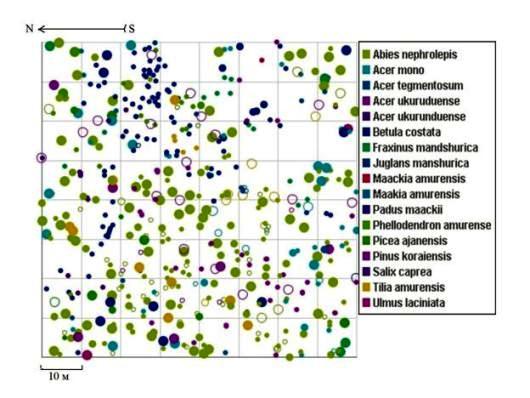
Рис. 113. Картосхема постоянной пробной площади пп 02-2002. Пихтово-кедровый лес на территории заповедника «Бастак» |
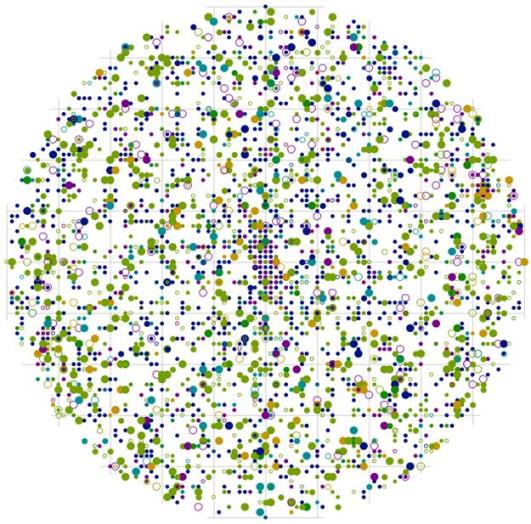
Рис. 114. Поле вероятности встречи деревьев в пределах фитогенного поля среднестатистической особи в пихтово-кедровом лесу (пп 02-2002) в заповеднике «Бастак» |

Рис. 115. Поле вероятности встречи деревьев в группе угнетенных пределах фитогенного поля среднестатистической особи этого класса толщины в пихтово-кедровом лесу (пп 02-2002) в заповеднике «Бастак» |
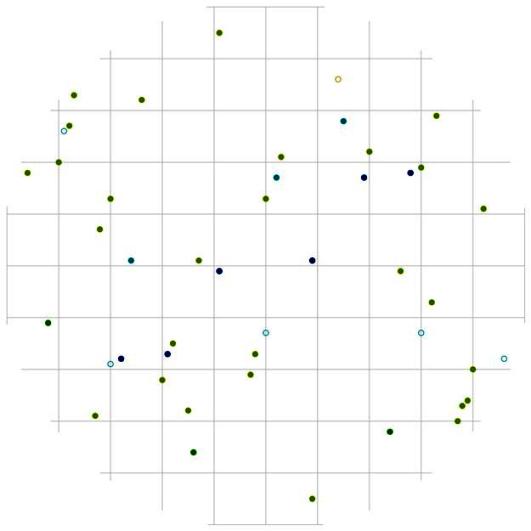
Рис. 116. Поле вероятности встречи деревьев в группе индетерминантных в пределах фитогенного поля среднестатистической особи этого класса толщины в пихтово-кедровом лесу (пп 02-2002) в заповеднике «Бастак» |
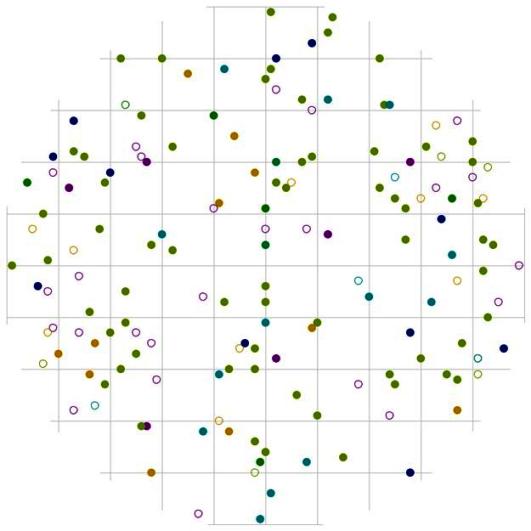
Рис. 117. Поле вероятности встречи деревьев в группе господствующих пределах фитогенного поля среднестатистической особи этого класса толщины в пихтово-кедровом лесу (пп 02-2002) в заповеднике «Бастак» |
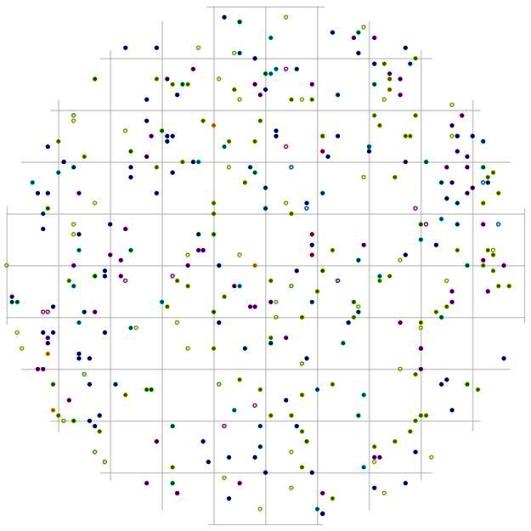
Рис. 118. Поле вероятности встречи деревьев группы угнетенных в пределах фитогенного поля среднестатистической особи господствующего класса толщины в пихтово-кедровом лесу (пп 02-2002) в заповеднике «Бастак» |
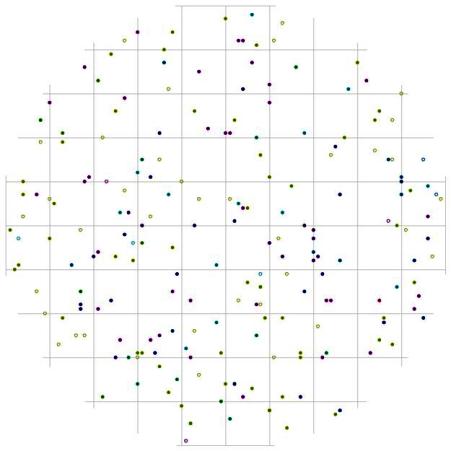
Рис. 119. Поле вероятности встречи деревьев группы угнетенных в пределах фитогенного поля среднестатистической особи индетерминантного класса толщины в пихтово-кедровом лесу (пп 02-2002) в заповеднике «Бастак» |
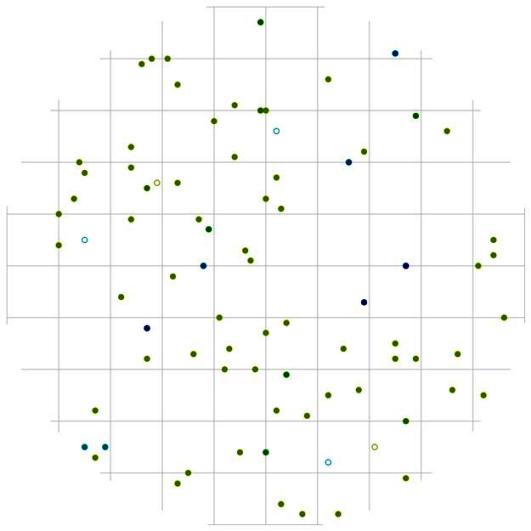
Рис. 120. Поле вероятности встречи деревьев группы индетерминантных в пределах фитогенного поля среднестатистической особи господствующего класса толщины в пихтово-кедровом лесу (пп 02-2002) в заповеднике «Бастак» |

Фото 20. Пихтовый с примесью кедра сибирского лес из пихты белокорой в Сихоте-Алинском заповеднике |
Таким образом, характер пространственного размещения разных ценотических групп по толщине в рассмотренных нами древостоях достаточно сложен и зависит от многих факторов, например, от внешних условий, в которых произрастает растительное сообщество. К ним относится положение (высота) над уровнем моря, крутизна склона и его ориентация, изначальная неоднородность микрорельефа, определяющая при формировании фитоценоза первичную мозаичность (Проскуряков, 1971. 1973). Анализ полученных данных говорит о том, что выработка социальной структуры древостоя в процессе самоорганизации имеет очень сложный характер, и проходит она в каждом древостое индивидуально. Полученные результаты подтверждают мнение многих авторов, приведенное в начале этого раздела о том, что простая классификация особенностей характера пространственной структуры древостоя по типам размещения не может вскрыть всех механизмов процесса выработки упорядоченности функциональных элементов в процессе самоорганизации древостоя. Даже при общей схожести картины распределения деревьев по ценотическим группам, в пространственном отношении картина размещения деревьев разных групп намного сложнее, каждый древостой характеризуется своим пространственным рисунком, который гармонирует со средой обитания и социальной структурой сообщества. Для каждого конкретного древостоя будет характерен свой пространственный ритм, который будет зависеть от первичной неоднородности среды и микрорельефа, а также от видов, составляющих данный ценоз, численности и количества ценотических групп. И проявляться, очевидно, он будет в наличии предпочитаемых и избегаемых расстояний и направлений в размещении деревьев как внутри групп, так и между особями разных ценотических классов.
Как показали наши данные, в древостоях с невыработанной ценотической структурой, находящихся на стадии восстановления или произрастающих в экстремальных условиях, ведущим фактором являются внешние условия и степень нарушенности не только самого древесного ценоза, но и субстрата, на котором данный ценоз произрастает. Именно внешний природный фактор чаще всего задает пространственный рисунок таких фитоценозов, а в отдельных случаях и сам человек (при искусственной посадке деревьев, после рубок или пожаров). Однако, как было показано выше, в процессе динамики древостоя происходит выработка социальной структуры древостоя в результате образования функциональных элементов системы. Растения, относящиеся к разным ценотическим группам и разным породам, начинают задавать свой, свойственный данному сообществу пространственный ритм (структуру размещения особей).
Одной из общих черт всех рассмотренных нами древостоев, независимо от степени хаотичности или, наоборот, устойчивости внутренней структуры, является неслучайный характер размещения особей разных пород деревьев и разного социального положения. Это подтверждается исследованиями и других авторов, указывающих на факт неслучайности произрастания разных видов растений в сообществах относительно друг друга (Галанин, 1989), а экспериментально доказанным фактом лучшего роста растений при совместном произрастании, если эти растения относятся к разным ценотическим группам (Плотников, 1974; Костенчук, 1985, и др.)