УДК 581.52
Высотная дифференциация растительности Сохондинского биосферного заповедника
Беликович А.В., Галанин А.В., Галанина И.А., Долгалева Л.М.*
Ботанический сад-институт ДВО РАН, г. Владивосток
* Институт технологии и бизнеса
Altitudinal differentiation of vegetation in Sokhondo Biosphere Reserve
Belikovich А .V., Galanin А .V., Galanina I.A., Dolgalyeva L. М .
Botanic Garden Institute of the Far-Eastern Branch of the Russian Academy of Science (BGI FEB RAS), Vladivostok
* Institute for Technology & Business , Nakhodka
Ключевые слова: растительность, высотные пояса, Сохондинский заповедник, Хентей-Чикойское нагорье, гора Сохондо
Altitudinal differentiation is one of factors of vegetation biodiversity in mountain areas. The authors describe vegetation belts in Khentei Mts and notice that a boundary between forest and steppe in Southern Transbaikalia constantly moves depending on climate cycles. In present times, in connection with the climate aridization cycle, more and more larch forests burn and the steppe vegetation element activates. In the period swith humid climate, prairieis replaced by forest, forest advances on a prairie.The results of vegetation monitoring on relevees in the Sokhondo Biosphere Reserve prove vegetation xerophitization in Souhern Transbaikalia and Northern Mongolia for the last hundred years. The upper tree line in the mountains is not stable; by observations of Nikota Sokolov in Sokhondo Mt in the middle 18 th century (250 years ago), climate here was more humid and tree line was 150 m above the present one. Special research proved that we have 600–700yr climate cycle in Dahuria Now the humid epoch is replaced by the arid period. Humidity maximum also took place in 11–12 th centuries, but the 13 th century was marked by climate aridization.
Введение
Высотная поясность или высотная зональность – это закономерная смена природных условий и ландшафтов в горах по мере возрастания абсолютной высоты (высоты над уровнем моря) (Википедия: http://ru.wikipedia.org/wiki/ ). Высотный пояс, высотная ландшафтная зона – единица высотно-зонального расчленения ландшафтов в горах. Высотный пояс образует полосу по природным условиям, часто прерывистую. Высотная поясность объясняется изменением климата с высотой: на 1 км подъёма температура воздуха снижается в среднем на 6°C, уменьшается давление воздуха, его запылённость, возрастает интенсивность солнечной радиации, до высоты 2–3 км увеличивается облачность и количество осадков. Между широтными зонами растительности и высотными поясами есть частичное сходство в климатических особенностях, в размещении растительности и почв. Но многим поясам невозможно найти полные широтные аналоги. Каждой широтной зоне свойствен свой тип высотной поясности, свой поясной ряд, характеризуемый числом поясов, их последовательностью и высотными границами. Полный спектр высотной поясности можно наблюдать в крупных горных массивах экваториальных и тропических широт (Анды, Гималаи). Самый нижний пояс в горной системе является продолжением той широтной зоны , которая расположена у подножия. Количество высотных поясов в каждой горной системе и их положение в основных чертах определяются широтой места и положением территории по отношению к морям и океанам. По мере продвижения с севера на юг высотное положение природных поясов в горах и их набор постепенно увеличиваются. Например, на Северном Урале леса поднимаются по склонам до высоты 700–800 м, на Южном – до 1000–1100 м, а на Кавказе – до 1800–2000 м. (Природные зоны и высотная поясность: http://www.ecosystema.ru/08nature/world/geoussr/3-1-1.htm)
Особи растений, составляющие растительный покров, неодинаковы, и неодинаковость растительного покрова бывает двоякого рода. С одной стороны, особи растений образуют генетические системы, называемые видами, в которых поколения особей связаны между собой потоками генетической информации. С другой стороны, расселяясь в пределах ландшафта, особи вступают в сложные взаимоотношения как с самим ландшафтом (средой обитания), так и друг с другом. Системы, возникающие в результате этого взаимодействия – ценоячейки, синузии, контурфитоценозы, микро- и мезокомбинации – и составляют растительность. Таким образом, растительность – это аспект социальной жизни растений.
Одним из факторов биоразнообразия растительного покрова в горных районах является его высотная поясность. Известно, что с высотой по мере подъема на каждые 100 метров климат в горах меняется так, как если бы мы переместились вдоль меридиана на несколько сотен километров. С высотой, так же как и с географической широтой, изменяется температура воздуха, количество осадков, продолжительность сезонов года. Однако высотная поясность растительности от поясности широтной или широтной зональности отличается двумя существенными особенностями. Если с географической широтой изменяется продолжительность дня и ночи, то при изменении высоты местности этот параметр не изменяется.
Второе существенное отличие высотной поясности от широтной заключается в том, что высотные пояса на несколько порядков уже, чем широтные. В связи с этим растительность нижнего высотного пояса может глубоко проникать в растительность следующего за ним по высоте пояса. По уникальным элементам микрорельефа растительность степного пояса, например, может проникать в нижний и даже верхний лесной пояса. При этом сообщества и ценопопуляции видов нижних поясов в горах приходят в непосредственное соприкосновение, а порой и во взаимопроникновение в сообщества и ценопопуляции верхних поясов растительнсоти. Это увеличивает вероятность интрогрессивной гибридизации родственных видов, приводит к увеличению разнообразия видов в растительных сообществах, к ускорению процессов видообразования.
Проблема высотной дифференциации растительности издавна привлекала внимание исследователей. В Европе исследователями альпийской растительности в XVIII – XIX вв. была разработана схема высотной поясности и предложена система понятий и терминов (лесной, субальпийский, альпийский, нивальный пояса), которую впоследствии геоботаники применяли при описании высотной дифференциации растительности всех других горных систем: Кавказа, Памира, Тянь-Шаня, Алтая, Саян, Сихотэ-Алиня и т.д. Однако довольно скоро ученые заметили, что схемы высотной поясности в разных горных системах могут существенно отличаться друг от друга, и здесь основное отличие связано с континентальностью климата – с тем, находится ли горная система близко к океану, или она удалена от него. В регионах с континентальным климатом поясность существенно отличается от таковой в регионах с океаническим климатом (Куминова, 1960; Тюлина, 1976; Огуреева, 1980). Разной бывает высотная поясность в горах, расположенных на Севере, и в горах, расположенных на Юге: на Севере ряд нижних высотных поясов в горах редуцирован, а в высокой Арктике вообще есть только один пояс. И только на экваторе могут быть представлены все пояса от вечнозеленых тропических лесов до горных тундр и альпийских лугов, и даже нивальный пояс здесь может быть выражен. Границы одних и тех же поясов на Юге и на Севере проходят на разной высоте. Например, на Полярном Урале нижняя граница пояса горных тундр проходит на высоте 100–200 м н.у.м., на Восточном Алтае на высоте 2000 м н.у.м., а в Гималаях на высоте 4000 м н.у.м.
В континентальных регионах взаимопроникновение растительности разных высотных поясов в горах бывает выражено сильнее, чем в регионах с океаническим климатом. Некоторые геоботаники такие регионы выделяли даже как регионы с уникальной структурой растительного покрова, где невозможно провести четкую границу между широтными поясами растительности. Так, например, растительный покров бассейна Колымы Ю.П. Пармузиным (1979) был выделен как особая природная зона – зона тундролесья. Именно тундролесья, а не лесотундры, так как здесь на южных склонах гор растут по всему высотному профилю настоящие лиственичные леса III и II бонитета, а на северных склонах по всему профилю развиты тундровые сообщества и заросли кедрового стланика. По широте с севера на юг пояс колымского тундролесья простирается на 1,5–2 тысячи километров.
В Забайкалье и в Южной Сибири степная зона выражена слабо, хотя аридные (степные) сообщества встречаются здесь в полосе шириной до 1 тысячи километров. Такие ландшафты геоботаники часто называют экспозиционной лесостепью. Здесь на южных склонах на выпуклых элементах мезорельефа до высоты 1700 м н.у.м. развиты степи и даурские прерии, а на северных склонах до высоты 900–1000 м н.у.м. опускаются лиственничные и сосновые леса. Дно межгорных впадин лежит здесь на высоте 700–900 м н.у.м. Провести границу между высотными поясами в таких регионах непросто.
Проблема усложняется еще и тем, что циклические вековые колебания климата приводят к смещению границ высотных поясов во времени. При аридизации климата степная растительность в горных районах начинает теснить лесную вплоть до того, что степной пояс входит в соприкосновение с горнотундровым. При гумидизации, напротив, активизируется лесная растительность, степи отступают вниз, а тундры отступают вверх. Во второй половине XX в. наука накопила достаточно фактов, чтобы оценить масштабы изменения высотной поясности в течение 600–700-летнего климатического цикла (Галанин, Беликович, 2002). Основная черта растительности южного Забайкалья и Сохондинского биосферного заповедника как его части – это динамичность и склонность к быстрой трансформации. Ритмичность природных процессов, вызванная как сезонными, так и тектоническими и гидрологическими колебаниями, способствует быстрой смене экосистем и растительных сообществ.
Прирусловые (пойменные) леса чаще рассматривались как азональные вне высотной поясности в связи со сложностью и многообразием этих лесов и неясно выраженной структуры. Исследования показали, что такие леса вдоль ручьев и рек высоких порядков также находятся в четкой корреляции с высотными отметками.
• Подгольцовый (1900–2100 м): в этой части прирусловые сообщества образуют узкие ленты пересыхающих ручьев с кедровыми редколесьями и кедрово-стланниковыми зарослями. Типичные черты гольцовой растительности ослабляются включением кустарников: Betula rotundifolia, Ribes nigra, Sorbaria sorbifolia.
• Верхний лесной (1600–1800 м): горно-таежные кедровые и кедрово-лиственничные леса. Широко представлены кедровники и лиственничники с кедром бруснично-зеленомышные, рододендроново-брусничные, баданово-брусничные. Для ручьев с подрусловым течением характерны голубичники и ивняки из Salixdivaricata и S . jenisseensis , протянутые узкой полосой вдоль русла. Междуречья обычно заняты довольно большим лиственничным редколесьем и ерниками. Характерен мохово-лишайниковый покров. По вогнутым долинам ручьев распространены высокобонитетные мохово-брусничные лиственничники.
• Нижний лесной (1300–1600 м н.у.м.): лиственничные, березо-лиственничные и сосновые леса до нижней границы леса на границе с лесостепями. По крупным рекам (Агуца, Агуцакан) обычны тополевые леса из Populus suaveolens. По расширенным долинам распространены ерники из Salix bebbiana, S . divaricata, Betula fruticosa, B. fusca. Аллювиальные галечно-песчаные поймы заняты Salix triandra, Salix viminalis, порой образующие непроходимые заросли.
• Степной пояс (800–1300 м н.у.м.) – тополевые, черемухово-яблоневые, сосновые, ивовые, березовые леса. Особенностью данного пояса является широкое распространение ивняков, протяженных вдоль всего русла широкой (до 100 м) полосой. Сосняки и березняки располагаются на высоких террасах, тополевые леса – зрелые, высокобонитетные, с большой степенью участия кустарников. Характерно близкое примыкание степей к речной пойме, слабая дифференциация лесной и степной флоры.
Высотное размещение прирусловых лесов отличают некоторые особенности, связанные с инверсией воздуха в ложбинах, микроклиматическими и орографическими особенностями местностей.
Целью данной статьи является описание и анализ особенностей высотной поясности растительности Сохондинского биосферного заповедника, расположенного в центральной части Хентей-Чикойского нагорья на границе Забайкальского края и Монголии (фото 1, 2).
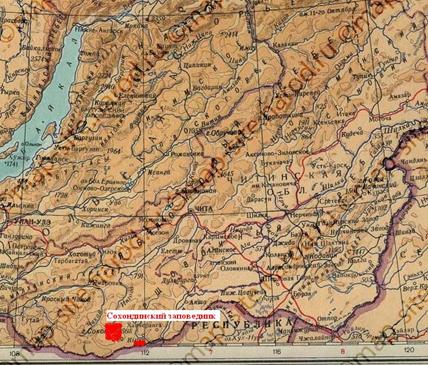
Фото 1. Положение Сохондинского биосферного заповедника и заказника «Горная степь» на географической карте
|
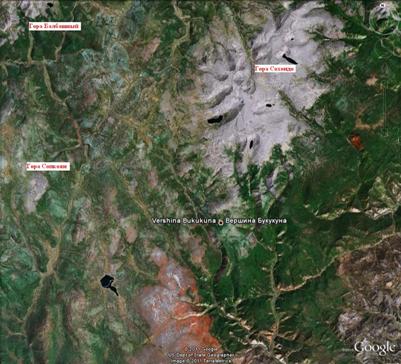
Фото 2. Космоснимок центральной части Сохондинского биосферного заповедника
|
Материалы и методы
Сохондинский биосферный заповедник является биосферный резерват ЮНЕСКО с 1985 г. Заповедник расположен на юге Забайкальского края. Его общая площадь 210988 га, а площадь охранной зоны 36060 га. (Сохондинский биосферный заповедник: http://www.kulttur.com/index).
Заповедник занимает наиболее возвышенную часть Хэнтей-Чикойского нагорья с гольцовым массивом Сохондо (фото 3). Горный массив Сохондо протянулся по периферии Хэнтей-Чикойского нагорья с юго-запада на северо-восток почти на 20 км при ширине до 14 км. Голец имеет две вершины – Большой Сохондо высотой 2505 м н.у.м. и Малый (2404 м), перевал между ними расположен на высоте 2000 м. Юго-западная часть массива носит название Цаган-Ула. С запада примыкает Джермалтай-Ингодинская депрессия. Характерная особенность указанного массива – изолированность друг от друга отдельных горных вершин, разделенных довольно глубокими котловинами и долинами рек. Кроме Сохондо, в пределах заповедника есть гольцы Сопкоян, Балбасный, небольшой водораздельный массив в истоках Киркуна и Буречи.
Прилегающая с юго-востока Алтан-Кыринская котловина находится на высоте 900–1000 м, так что перепад высот довольно значителен (до 1,5 км). Очень характерны для Сохондо гольцовые террасы, сформированные различными обломками горных пород. На плато имеются своеобразные каменные колодцы. Склоны гор различной крутизны, но наиболее типичны в среднем 15–20 градусов. В зимнее время на крутых склонах террас, каров и троговых долин могут появляться снежные лавины, образуя при сходе своеобразные желоба с лавинными конусами в устьях. Благодаря морозному выветриванию, а также под влиянием местных подвижек земной поверхности, каменные россыпи медленно, но почти непрерывно движутся вниз по склонам. Долины рек, возникшие под влиянием ледниковой эрозии хорошо разработаны, становясь круче у подножия гор, где приобретают V-образную форму.

Фото 3. Вершина горы Сохондо. Вид из верхнего лесного пояса. Фото А.В. Беликович |
Климат отличается резкой континентальностью. Характерна сухая и малоснежная зима антициклонального типа со среднемесячными температурами января в пределах минус 22–28 град. В зависимости от высот (минимумы от -40 до -55 градусов). По данным гидрометеостанции «Букукун» средняя многолетняя температура января -19,8 град. С. Продолжительность устойчивого снежного покрова 130–145 дней. Глубина снега колеблется от 70–100 см в кедровниках верхней полосы лесного пояса до 5–15 см в смешанных лесах нижней полосы лесов. В лесостепи и по долинам крупных рек сплошной снежный покров, как правило, не образуется. Продолжительность солнечного сияния достигает 3000 часов в год. Во второй половине зимы и в начале весны преобладает ясная и тихая морозная погода. Весна сухая и ветреная. Погода в этот период неустойчива: то сухо и тепло, то возникают снежные бураны. Лето короткое; в июне возможны снегопады и заморозки. Для второй половины лета характерны затяжные муссонные дожди, вызывающие паводки на горных реках. Самый жаркий месяц – июль (среднегодовая температура +14,7 град. С). Осень теплая и сухая, первые заморозки – в начале сентября. Среднегодовое количество осадков для степного пояса 430 мм, среднегодовая температура воздуха 1,9 град. С. |
Гидрологическая сеть заповедника довольно густая и характеризуется множеством рек и озёр. Большинство рек берут своё начало с гольцовых массивов (Сохондо, Сопкоян, Балбасный) и их отрогов и имеют ледниковое питание. В истоках реки узкие, иногда вода струится под камнями и глыбами, не выходя на поверхность. Спускаясь с гор, реки образуют широкие, часто заболоченные долины. Питание мелких рек преимущественно дождевое, роль грунтовых вод незначительна. Поскольку мерзлота образует водонепроницаемый слой, летние осадки стекают в реки, вызывая паводки. Холодные малоснежные зимы способствуют распространению наледей, а реки часто промерзают до дна. На северных склонах наледи иногда сохраняются до августа. Имеется в заповеднике и множество минеральных источников (аршанов), активно посещаемых животными для восполнения недостатка солей в организме. Наиболее примечательные – Ендинский (кл. Кислый), Агуцаканский и Ингодинский (кл. Аршан), которые относятся к гидрокарбонатному-хлоридо-натриевому и сернисто-водородному классам.
Мелкие моренные озера лежат цепью в районе мирового водораздела рек Тихого и Северного Ледовитого океанов в центре заповедника близ гольца Сохондо. Более крупные озера, такие как Нарья (площадь водной поверхности 120 га), Угдыри, богаты растительностью и привлекают к себе внимание водоплавающих (поганки, различные утки, чернозобая гагара, кулики) и сохатых. Озера ледникового происхождения находятся в обрамлении стенок каров гольцов и выглядят весьма живописно. Они довольно глубоки (порядка 20 м и более). Из-за низких температур планктон и водоросли в них почти не развиваются, поэтому прозрачная вода в них приятного голубоватого оттенка. В знаменитом Букукунском озере сложилась замкнутая популяция ленка, которая во многом отличается от речной формы, т.к. истоки реки Букукун загромождены каменистыми россыпями.
На сухостепных участках заповедника формируются горные каштановые почвы, наиболее характерные для пологих склонов. Долинам рек свойственно большое разнообразие почвенных типов. По днищам падей формируются лугово-черноземные почвы, а на повышенных участках – дерново-луговые. Под лугами и болотами образуются наиболее гидроморфные торфянисто-глеевые почвы. Наиболее распространенными типами лесных почв являются мерзлотно-таежные и горные мерзлотные дерново-таежные, развитые под лиственничниками и мшистыми лесами на склонах северных экспозиций. При этом дерново-таежные почвы формируются в более благоприятных гидротермических условиях по нижним частям склонов. Второй характерный тип лесных почв (после дерново-таежных) – горные серые лесные почвы, развитые под травяными березняками или иными осветленными лесами с многоярусным травяным покровом. На хорошо дренированных участках фрагментарно встречаются горные подзолистые и дерново-подзолистые почвы.
В основу статьи положены материалы, собранные авторами на протяжении длительного времени – с 1982 по 2010 гг. – во время экспедиций и стационарных исследований. В сборе материала участвовали все авторы. А.В. Галанин и А.В. Беликович изучали флору и растительность заповедника с 1982 по 2006 гг. И.А. Галанина – в 2001 и 2002 гг. Л.М. Долгалева в 2004–2008 гг. Суммарная длина пешеходных маршрутов в заповеднике и его окрестностях всех авторов составила более 500 км, при этом было составлено более 1000 маршрутных геоботанических описаний и более 50 описаний постоянных пробных площадей размером 50х50м. Описания и постоянные пробные площади располагались вдоль высотных профилей от дна Алтано-Кыринской и Ононской котловин до вершин гор Сохондо, Балбашный и Сопкоян.
С 2002 по 2004 г. ботанические исследования осуществлялись в соответствии с договором о научном сотрудничестве между Ботаническим садом-институтом ДВО РАН и Сохондинским биосферным заповедником, а начиная с 2004 г. наши работы по изучению растительного мира Даурии осуществлялись в рамках Программы комплексных исследований в бассейне реки Амур и по гранту ДВО РАН «Флора Даурии». В экспедициях-исследованиях, кроме авторов статьи, активно участвовали: Е.Н. Роенко, Л.С. Яковченко, И.В. Козырь, В.А. Галанин, а также д.б.н. С.Б. Гончарова, д.б.н. И.Н. Сафронова, д.б.н. О.М. Афонина, к.б.н. Е.О. Головина. Исследование было бы невозможно без содействия директора Сохондинского заповедника В.И. Яшнова, зам. директора по научной работе к.б.н. Е.Э. Малкова, зам. директора по охране А.В. Малышева, научного сотрудника И. Белова, инспекторов Г. Слесаренко и И. Маврина, водителя В.П. Пляскина. В обработке гербарного материала участвовали И.Ю. Орловская и Р.Г. Кирлан. Мы выражаем им свою признательность.
При описании растительности мы использовали традиционные методы геоботаники (Шенников, 1964; Быков, 1970; Воронов, 1973): метод пробных площадей, метод детального картирования микрогруппировок, картирование древостоя, проводили таксацию древостоя и ревизию пробных площадей, заложенных при лесоустройстве заповедника в 1978 г. Полученные данные сводили в таблицы и компьютерную базу. При обработке использовали программы статистической обработки данных, космоснимок территории заповедника взят с портала Google Earth.
Результаты
По нашим данным флора сосудистых растений Сохондинского заповедника и его охранной зоны насчитывает 913 видов из 313 родов и 75 семейств. Ведущими семействами в порядке убывания в них количества видов являются: Compositae, Poacea, Ranunculaeceae, Rosaceae, Fabaceae, Liliaceae, Scrophulariaceae, Brassicaceae, Caryophillaceae, Salicaceae, Apiaceae, Polygonaceae. Ведущими родами во флоре заповедника являются: Carex, Salix, Potentilla, Poa, Allium, Gentiana, Artemisia, Viola, Oxytropis, Ranunculus, Betula, Calamagrostis, Juncus Astragalus (Беликович, Галанин, 2002).
По данным лесоустройства 1978 г. площадь лесов составляет 83% от территории заповедника. Преобладающей породой является лиственница (Larix dahurica) – 55% лесопокрытой площади, затем кедр (Pinus sibirica) – 28,3%, далее идет кедровый стланик (Pinus pumila) – 5,3%. Сосна обыкновенная (Pinus sylvestris) и лиственные породы занимают в сумме около 5,6%. Нелесная площадь представлена тундрами, каменными россыпями, покрытыми накипными лишайниками, болотами, и составляет 17% от всей площади. Небольшие участки степей-прерий встречаются по долинам рек, примыкающих к Алтано-Кыринской котловине и на склонах гор южной экспозиции.
В связи с сильно пересеченным горным рельефом создаются благоприятные условия для встречных миграций и контактов высокогорных и низкогорных видов. Поэтому для растительности заповедника характерна гетерогенность видового состава, проявляющаяся в совместном произрастании растений различных экологических групп. Например, в степном сообществе наряду со степными могут произрастать лугово-лесные и даже лугово-болотные виды. То же самое можно сказать о луговой растительности. В большинстве случаев это молодые растительные формации, возникшие на месте ивово-ерниковых зарослей и различных по составу лесов, уничтоженных рубками или пожарами. В составе луговой растительности следует отметить наличие «сниженных альпийцев», обычными местами обитания которых являются альпийские лужайки, редколесья и кустарниковые заросли высокогорий около ледников или тающих снегов.
Лиственничные леса занимают склоны северных экспозиций, заболоченные террасы и речные долины. Они растут на маломощных почвах, играя водо-охранную роль. Сосновые леса (Pinus sylvestris) занимают относительно теплые, хорошо освещенные склоны южных экспозиций в нижней части лесного пояса. Это чистые или смешанные леса с ярусом из Betula platyphlla, густым подлеском из Rhododendron dauricum, при участии видов родов Spiraea и Cotoneaster. В лесах нижнего лесного пояса хорошо развит травяной покров, под полог сосновых и лиственничных лесов заходят многие виды из дауских прерий и степей. Сосновые леса тяготеют к выходам гранитов, песчаникам и песчаным субстратам. Лиственничные – к глинистым сланцам и продуктам их выветривания. Темнохвойные леса представлены в первую очередь кедровниками разных типов (Pinus sibirica) и кедрово-лиственничными лесами, среди которых выделяются наиболее продуктивные, приуроченные к границе верхнего и нижнего лесных поясов. Древостой здесь обычно состоит из двух хвойных пород.
В верхнем лесном поясе (фото 4, 5, 6) преобладают кедрово-лиственничные и лиственнично-кедровые кустарничково-зеленомошные леса с хорошо развитым ярусом из багульника (Ledum palustre), голубики (Vaccinium uliginosum) и брусники (Rhodococcum vitis - idaea). Кедровники чернично-брусничные встречаются в верхнем лесном и субальпийском поясах. Наземный покров в них низкий, распределен пятнами, флористически бедный, в нем преобладают Vaccinium murtillus и Rhodococcum vitis - idaea ; зеленые мхи покрывают до 80% поверхности почвы, кустистые лишайники – до 5%.

Фото 4. Верхний лесной пояс в верховьях реки Букукун. Фото А.В. Галанина
|

Фото 5. Кедрово-лиственничный багульниковый лес в верхнем лесном поясе. Фото А.В. Галанина |

Фото 6. Лиственнично-кедровый баданово-кустарничковый лес с подлеском из кедрового стланика на границе верхнего лесного и субальпийского поясов. Фото Л.М. Долгалевой
|
Леса из Betula platyphilla обычно развиваются на месте сгоревших или вырубленных хвойных лесов в нижних частях склонов гор и на террасах. В долинах больших рек (Агуца, Буреча, Букукун) растут разнообразные старые тополевники (Populus suaveolens), преимущественно разнотравно-злаковые с густым и сложным наземным покровом. Осиновые леса (Populus tremula) встречаются только на крутых прогреваемых склонах гор южной экспозиции. Осина в заповеднике ведет себя как типичный ксерофит, осинники на склонах обычно обрамляют пятна даурской прерии.
Степная растительность (фото 7) на территории заповедника встречается по долинам рек Агуцы, Агуцакана, Букукуна, Енды, Киркуна, Буречи фрагментами на склонах южной и юго-восточной экспозиции до высоты 1600–1700 м н.у.м., а иногда и значительно выше – до 2000 м н.у.м., где смыкается с высокогорными тундрами (гора Сопкоян). Однако площадь степных сообществ в заповеднике невелика. Благодаря нашим настойчивым рекомендациям и позиции директора В.И. Яшнова в 2003 г. на восточном макросклоне Онон-Бальджинского хребта, обращенном в сторону реки Онон, вдоль российско-монгольской границы создан заказник «Горная степь» (фото 8), в котором хорошо представлена степная растительность. Заказник подчинен заповеднику и, по сути, является его составной частью. |

Фото 7. Ковыльная степь в долине реки Онон в окрестностях с. Дадал (Монголия). Фото И.Н. Сафроновой
|

Фото 8. Лесостепной пояс в заказнике «Горная степь» на восточном макросклоне Онон-Бальджинского хребта. Фото А.В. Галанина |

Фото 9. Луковая прерия в долине реки Онон в окрестностях с. Дадал (Монголия). Фото И.Н. Сафроновой
|

Фото 10. Бубенчиковая прерия в долине реки Онон в окрестностях с. Дадал (Монголия). Фото И.Н. Сафроновой
|

Фото 11. Харганат абрикосовый в заказнике «Горная степь». Фото А.В. Галанина
|
К криофильной (условно степной) растительности в Сохондинском заповеднике мы относим три основных типа: 1 – даурские прерии (фото 9, 10) (разнотравные, нителистниковые из Filifolium sibiricum), 2 – даурские степи (ковыльные, типчаковые и осочковые) и 3 – харганаты (абрикосовые, ильмовые, спирейные) (фото 11).
Непременным элементом растительности заповедника являются долинные степи, распространенные по плоским гривам речных террас. В травостое преобладают злаки.
Для высокогорий заповедника характерны дриадово-разнотравно-лишайниковые тундры, в которых обильны Dryas oxyodontha, Vaccinium uliginosum ssp. microphylla, небольшие фрагменты альпийских разнотравных и осоково-моховых лугов. Здесь господствуют мохово-лишайниковые тундры. Среди кустарников высокогорий господствует кедровый стланик (Pinus pumila). В отдельных случаях присутствуют Rhododendron аureum, Salix divaricata, Dushecia fruticosa, Betula nana ssp. rotundifolia и др. Наземный покров представлен зелеными мхами в прогалинах стланика и лишайниками под его пологом.
Изучая растительность Даурии, мы пришли к выводу, что растительный покров этого региона ботанически более своеобразен, чем это считалось до настоящего времени, что здесь, кроме известных типов растительности, исторически сложились два своеобразных типа, которые отсутствуют за пределами Даурии. Ранее один из этих типов растительных сообществ геоботаники классифицировали как кустарниковые степи, а другой – как разнотравные степи, хотя со степной растительностью они имеют весьма мало общего. |
Мы считаем, что главными характерными особенностями типа растительности являются объем, скорость и степень замкнутости биологического круговорота, обеспечиваемого растительными сообществами этого типа. Эти параметры биологического круговорота вещества в наибольшей степени зависят от характера жизненных форм растений, сообитающих друг с другом в конкретных растительных сообществах (Галанин, 1989; 1991; 2005). Именно поэтому характеризуя каждый тип растительности, геоботаник обязательно называет жизненную форму доминирующих растений. Луга – это сообщества травянистых многолетних мезофитов, чаще всего корневищных многолетников; степи – сообщества травянистых ксерофитов с доминированием плотнодерновинных злаков, короткокорневищных злаков, осок и луковичных эфемероидов; светлохвойные леса – сообщества хвойных листопадных деревьев (лиственницы); темнохвойные леса – сообщества хвойных зимнезеленых деревьев (ели, пихты, псевдотсуги и др.); листопадные широколиственные леса – сообщества широколиственных деревьев, сбрасывающих на зиму листву (дуб, липа, ясень, клен и др.). Каждый тип растительности имеет свою длинную историю, насчитывающую десятки, а иногда и сотни миллионов лет, в течение которой происходила коэволюция совместно обитающих видов растений и животных. Эта эволюция постоянно корректировалась определенными климатическими и экологическими условиями внешней среды, с которыми данный тип растительности находился в относительном биогеохимическом равновесии.
За многие миллионы лет в растительном покрове Земли выработались весьма устойчивые сочетания жизненных форм и видов живых организмов, обеспечивающие относительную биогеохимическую устойчивость ландшафтных экосистем, – для каждого типа элементарного ландшафта сформировался свой особый тип экосистем и свой особый тип растительности. В умеренной зоне при дефиците воды в течение почти всего вегетационного периода и при относительно хорошем увлажнении весной за счет тающего снега и дождей, не редких в это время года, устойчивым типом растительности являются степи, которые буквально расцветают за счет эфемероидов весной, но летом сильно усыхают. В степях запас фитомассы над поверхностью почвы в 7–8 раз меньше, чем запас фитомассы в корнях, луковицах и корневищах под землей (Cочава, 1964). Отмирающая фитомасса в степи разлагается медленно, поэтому здесь всегда имеется избыток органического вещества и в почве (черноземы) и на ее поверхности (прошлогодняя ветошь).
В умеренной зоне, но при отсутствии устойчивого дефицита воды в течение вегетационного периода, там, где сильно выраженный аллювиальный процесс не позволяет расти деревьям и высоким кустарникам, устойчивым типом растительности являются луга. Для лугов характерно значительно меньшее преобладание подземной фитомассы над надземной, процессы гниения здесь идут значительно интенсивнее, чем в степях. Черноземы под лугами не формируются. Сходным в степях и на лугах является то, что практически вся надземная фитомасса, нарастающая за вегетационный сезон, осенью отмирает, многолетними являются только подземные органы, из которых на следующее лето отрастает новая надземная фитомасса. В отличие от степей, на лугах разница в надземной и подземной фитомассе значительно меньше: в подземной сфере сосредоточено примерно две трети общего запаса фитомассы.
Для типа растительности верховое болото характерно еще более медленное, чем в степях, разложение отмершей фитомассы. Она буквально консервируется здесь на многие десятки и сотни тысяч лет, накапливаясь в виде торфов. Характерной жизненной формой для этого типа растительности являются сфагновые мхи – природные консерванты мертвой органики. Верховые болота образуются в условиях постоянного избыточного увлажнения в течение всего вегетационного периода. Болотные экосистемы характеризуются разомкнутым биологическим круговоротом вещества, чем напоминают экосистемы океана. Есть основание считать, что это очень древний тип экосистем (Галанин, 1989). Эволюция биосферы, по крайней мере, в мезозойскую и кайнозойскую эры, происходила в направлении формирования экосистем, работающих по замкнутому типу и в направлении постоянного ускорения оборота вещества в трофических цепях жизни.
Тип растительности темнохвойная тайга отличается тем, что здесь значительная фитомасса накапливается в стволах и корнях деревьев и на многие десятки лет выводится из ежегодного биологического круговорота. Хвоя здесь живет 3–4 (5) лет, что также приводит к существенному замедлению биологического круговорота вещества. По замедленности биологического круговорота темнохвойные леса ближе к степям, но по типу «складирования» органического вещества, исключенного из круговорота, они есть нечто совершенно иное. Склад исключенного из ежегодного круговорота вещества в этих экосистемах содержится не только в почве, но и в еще большей степени в самих растениях, в основном в стволах деревьев. Темнохвойная тайга устойчива в условиях бореального климата с достаточным в течение всего вегетационного периода увлажнением. Светлохвойная тайга от тайги темнохвойной отличается тем, что хвоя здесь отмирает ежегодно, поэтому биологический круговорот в экосистемах светлохвойной тайги интенсивнее, чем в экосистемах темнохвойной тайге. Интенсификации круговорота вещества в светлохвойной тайге способствует и то, что весной и в начале лета, пока хвоя на деревьях не отросла полностью, под осветленным пологом леса интенсивно развиваются кустарники и травянистые растения. Ежегодно отмирающая наземная фитомасса травянистых растений в светлохвойных лесах – это весьма существенный элемент ежегодного биологического круговорота вещества в экосистемах светлохвойной тайги. Светлохвойная тайга устойчива в условиях резко континентального бореального климата в районах, где зимой выпадает мало снега, а почва сильно промерзает так, что нередко образуется многолетняя мерзлота, а весна и начало лета отличаются крайней сухостью.
Но вернемся к Даурии. Для этого региона характерен очень своеобразный ритм погоды, настолько своеобразный, что здесь уместно говорить не о четырех, а о пяти сезонах года. Первая половина лета (с 20 апреля по 20 июля) отличается крайней сухостью, это особый сезон года – сухое лето. Вторая половина лета (с 20 июля по 10 сентября) – это период, когда количество выпадающих осадков значительное, вполне достаточное для роста и развития растений мезофитов. Это также особый сезон года – влажное лето. Подобный ритм погоды сохраняется в Ононской Даурии на протяжении многих сотен тысяч (а может быть и двух миллионов) лет благодаря тихоокеанскому муссону (Маркевич, 1990). За столь длительный период времени в здешних ландшафтах сформировалось, по крайней мере, два своеобразных типа растительных сообществ, аналогов которым нет в других регионах северной Евразии: это даурская прерия и харганат. Впервые сообщества ксерофитов и мезофитов Даурии как прерии классифицировал Р.В. Камелин (1987).
Даурская прерия представляет собой оригинальное сочетание настоящих травянистых ксерофитов, мезофитов и мезо-ксерофитов. В период сухого лета в экосистеме биогеохимически активны ксерофиты, рост и развитие которых осуществляется за счет запаса питательных веществ, накопленных в корневищах. В период влажного лета сильно активизируются мезофиты и мезоксерофиты. Смесь ксерофитов и мезофитов в одном сообществе на одном участке – характерная особенность даурской прерии. В июне эта прерия выглядит как сухая степь или даже полупустыня. Общий облик этой прерии в это время бурый, зелень едва пробивается и плохо видна среди прошлогодней ветоши. Зато в августе сухая даурская прерия превращается в зеленый цветущий луг. В табл. 1 приведен обобщенный список видов даурской прерии, составленный в результате анализа и обобщения нескольких десятков конкретных геоботанических описаний, составленных нами в пределах Ононской Даурии за весь период наших исследований, начиная с 1982 г.
Таблица 1
Общая характеристика видового состава нового типа растительности даурская прерия (лугостепь)
Мезофиты
|
Ксерофиты и мезоксерофиты
|
Обильны |
обильны |
Potentilla acervata
Potentilla tanacetifolia
Sanguisorba officinalis
Artemisia tanacetifolia
Galium verum |
Leontopodium conglobatum
Stellera chamaejasme
Bupleurum sibiricum
Filifolium sibiricum |
Обычны |
обычны |
Polygonum angustifolium
Pentaphylloides fruticosa
Potentilla fragarioides
Anemone crinita
Geranium vlassovianum
Astragalus dahuricus
Fragaria orientalis
Rhaponticum uniflora
Vicia cracca
Rhumex thyrsiflorus
Veronica longifolia
Delphinium grandiflorum |
Senecio amurensis
Heteropappus altaicus
Scabiosa comosa
Veronica dahurica
Artemisia sp. nova
Adenophora stenanthina
Stipa baicalense
Shizonepeta multifida
Plantago depressa
|
нередки |
нередки |
Lophanthus chinense
Halenia corniculata
Polygonum alopecuroides
Silene chamaerensis
Astragalus membranaceus
Campanula glomerata
Adenophora coronopifolia
Adenophora tricuspidata
Linaria acutiloba
Gentiana triflora
Lilium dahuricum
Myosotis caespitosa
Lilium pumilum
Filipendula palmata
Polygonatum odoratum
Ligularia fischeri
Vicia unijuga
Trifolium lupinaster
Sanguisoeba officinalis |
Potentilla anserinoides
Potentilla conferta
Echinops latifolius
Stipa sibirica
Goniolimon prostratum
Gypsophyla dahurica
Thalictrum petaloideum
Veronica linarifolia
Thymus dahuricus
Helictotrichon shellianum
Aster serpentimontanus
Sedum purpureum
Chamaerhodos erecta
Kobresia filifolia
Dianthus repens
Lespedeza hedysaroides
Rheum undulatum
Sedum aizoon
Pulsatilla turczaninowii
Polygonum divaricatum
Patrinia rupestris
Aster tataricus |
В ландшафте даурская прерия занимает высокие сухие участки надпойменных террас, сложенных галькой с примесью песка, а также щебнистые и мелкокаменистые склоны гор южной и юго-восточной экспозиций. Видовой состав даурской прерии, особенно состав доминирующих в ней видов растений, может изменяться в зависимости от крутизны склона, размера камней и щебня, содержания в субстрате мелкозема, характера материнской горной породы. То, какие виды растений в прерии будут доминировать, в значительной степени определяется и ценотическим фактором, а конкретнее тем, какие виды окажутся победителями в процессе многолетней конкуренции за воду и минеральные вещества. Нестабильность погодных условий от года к году и вековые колебания аридности-гумидности климата в Центральной Азии приводят к тому, что процессы конкуренции здесь никогда не успевают привести к вытеснению из растительных сообществ многих видов, поэтому сообщества остаются многовидовыми. Действительно, в сырые годы конкурентно способнее оказываются растения мезофиты, а в сухие – растения ксерофиты. Чем нестабильнее условия среды, тем большее количество видов может выжить в конкретном сообществе на конкретном участке. Столь высокое таксономическое разнообразие даурской прерии прежде всего говорит о том, что климатические и экологические условия здесь в высшей мере нестабильны на протяжении длительного времени.
Обычно принято считать, что чем больше в фитоценозе редких и малообильных видов, тем он устойчивее, тем больше его резистентность. Думаю, что это не так однозначно. В том случае, когда климатические условия изменяются циклически, в одном сообществе могут сосуществовать виды разного экологического склада: мезофиты и ксерофиты – в степях-прериях Даурии, мезофиты и криофиты – в луговинных тундрах Южной Чукотки (Беликович, 2001). Такой ксерофитно-мезофитный маятник раскачивается в Даурии, по крайней мере, в течение нескольких десятков тысяч лет.
Еще более своеобразным типом растительности, также специфичным для Ононской Даурии, являются сообщества с доминированием древесных растений (Беликович, Галанин, 2005) – очень низкрослых деревьев (Armeniaca sibirica, Ulmus pumila, U. macrocarpa, Malus pallasiana) и кустарников (Spiraea aquilegifolia, Ribes diacanthum, Pentaphylloides parviflora, и др.) Эти сообщества в ландшафте в пределах степного и лесостепного поясов занимают крутые каменистые и скалистые склоны южной и юго-восточной экспозиций, явно тяготея к выходам глинистых сланцев и известняков и избегая граниты и гранито-гнейсы. Местные жители российской части Даурии этот тип растительности называют абрикосниками, жители монгольской части Даурии – харганатом (фото 12). Ботаники чаще всего относят их к кустарниковыми степям (Дулепова, 1979). Мы считаем такие сообщества своеобразным типом растительности и предлагаем сохранить за ним название харганат (Беликович, Галанин, 2005).

Фото 12. Харганат ильмовый в заказнике «Горная степь». Фото А.В. Галанина |
Высота деревьев и кустарников харганата обычно не превышает 1,5–2 м, они часто страдают от пожаров, однако отлично возобновляются и быстро отрастают в виде корневой поросли. У ильма крупноплодного имеется превосходное приспособление переносить пожары. На его веточках и стволиках в условиях частых пожаров образуются крылатые пробковые выросты, предохраняющие меристематические ткани от высокой температуры. При высоте деревцев и кустов в полтора метра их возраст может, по-видимому, достигать нескольких сотен лет. Травянистые растения в харганате не образуют сплошного покрова. Чаще всего они встречаются в виде отдельных куртин с проективным покрытием оснований менее 30–40%.
Интересной особенностью харганата является то, что при резком снижении пресса пожаров он превращается в низкорослый (3–4 м) разреженный ильмовый лес с подлеском из сибирского абрикоса и густым ярусом кустарников. Высота деревьев в таком лесу 3–4(5) м, сомкнутость крон до 30%, а толщина деревьев ильма может достигать 30 см в диаметре. Такие ильмовые рощицы мы неоднократно встречали у основания каменистых склонов, где иногда они занимали площади в несколько гектар. Вообще ильмы в Ононской даурии не редкость, хотя настоящие древостои они здесь образуют редко. Надо полагать, что в более гумидные, чем современная, эпохи ильмовые леса были вполне обычным элементом здешнего ландшафта.
|
В верхней части склонов южной и юго-восточной экспозиций в гумидные эпохи в Сохондинском заповеднике, по всей вероятности, росли дуб монгольский (Quercus mongolica) и береза даурская, или черная (Betula dahurica), которые в настоящее время как реликты встречаются в восточной части Даурии Аргунской.
Мы считаем, что оба новых типа растительности – даурская прерия и харганат – требуют самого тщательного изучения. Замечательно то, что в 2003 г., благодаря присоединению к Сохондинскому биосферному заповеднику большого участка в лесо-степном поясе на восточном макросклоне Онон-Бальджинского хребта, в заповедном фонде оказались и даурская прерия (фото 9, 10), и харганат (фото 12).
Особенностью высотного распределения растительности в Сохондинском заповеднике является то, что на южных и северных склонах здесь развиваются очень контрастные растительные сообщества: на северных – леса, а на южных – степи, даурские прерии и харганаты. Даурские прерии и харганаты порой достигают верхнего лесного и даже гольцового поясов, так что точно и однозначно назвать высотные пределы каждого пояса невозможно. В свою очередь, кустарниковая и кустарничковая растительность по долинам рек и ручьев из гольцового пояса нередко спускается в нижний лесной и даже в лесостепной пояса. Такие высотные инверсии типов растительности обусловлены резкой континентальностью климата Даурии, влиянием тихоокеанского муссона во второй половине лета и широким распространением в горах Хэнтей-Чикойского нагорья сплошной в альпийском поясе гор и островной в лесном и лесостепном поясах многолетней мерзлоты. Малоснежные и бесснежные зимы, сухая возгонка снега под воздействием ультрафиолетового излучения, доля которого в солнечном спектре на высотах 700–2500 м н.у.м. весьма значительна, крайняя сухость воздуха зимой, весной и в начале лета создают в ландшафтах Хэнтея и Ононской Даурии очень контрастный и разнообразный спектр местообитаний и высокое разнообразие растительных сообществ.
Обработка и анализ собранного нами материала, а также анализ литературы по растительности Даурии и сопредельных территорий, позволили выявить закономерности высотной поясности и существенно уточнить схему высотной поясности с учетом особенностей климата и характера подстилающих горных пород.
В пределах Сохондинского заповедника и его охранной зоны нами выделяются 6 высотных поясов: степной, лесостепной, нижний лесной восточносибирских лиственничных лесов, верхний лесной кедрово-лиственничных лесов, подгольцовый (субальпийский) или кедрово-лиственничных редин и кедрового стланика, гольцовый (альпийский) дриадово-разнотравных и эпифитно-лишайниковых тундр и альпийских лужаек.
Степной пояс здесь занимает высоты до 600–700 м н.у.м. В пределах этого пояса встречаются ковыльные и разнотравно-злаковые степи, даурские прерии, разнотравные и разнотравно-злаковые (полевицевые) луга. По берегам ручьев и рек здесь встречаются ивняки. Степной пояс здесь занимает высоты до 600–700 м н.у.м. В пределах этого пояса встречаются ковыльные и разнотравно-злаковые степи, даурские прерии, разнотравные и разнотравно-злаковые (полевицевые) луга. По берегам ручьев и рек здесь встречаются кустарниковые ивняки и ивовые леса, со всеми характеристиками лесного сообщества – ярусностью, сменой доминатов, дифференцированной структурой древостоя. Наиболее явные признаки таких лесов наблюдаются в долинах заповедной территории, не подверженной длительное время пожарам. В степной зоне обычны вдоль рек и разреженные березняки. Наиболее широко распространены разнотравные сообщества. В этих сообществах содоминируют Galium verum, Bupleurum scorzonerifolium, Lespedeza hedysaroides, Filifolium sibiricum, Potentilla acervata, P. tanacetifolia, Scutellaria baicalensis, Schizonepeta multifida, Adenophora gmelinii, Gypsophila davurica. Как правило, в сложении травостоя участвуют злаки – Stipa baicalensis, Achnatherum sibircum, Poa attenuata, Carex pediformis. Нителистниковые (Filifolium sibiricum) сообщества характерны для склонов средней крутизны (с углом наклона примерно 20?–40?), с щебнистой, каменисто-щебнистой почвой, с небольшими выходами пород. На участках с каменисто-щебнистой почвой (обычно в привершинных частях склонов и иногда на вершинах) обилие разнотравья незначительно, заметную роль в сообществах играют Festuca lenensis и Artemisia monostachya. Для этих сообществ характерны такие виды разнотравья, как Phlojodicarpus sibirica, Bupleurum bicaule, Eremogone capillaris, Pulsatilla tenuiloba; из кустарников – Pentaphylloides parvifolia. Холоднополынные (Artemisia frigida) сообщества развиты в нижних частях склонов и на шлейфах, до недавнего времени подвергавшихся выпасу скота.
Лесостепной пояс (фото 13), или, пояс экспозиционной лесостепи занимает районы низкогорий с высотами до 1400 м н.у.м. Здесь наблюдается четкая приуроченность сосновых и лиственнично-березовых лесов к склонам северных экспозиций, а даурских прерий, разнотравно-злаковых и ковыльных степей – к склонам южных экспозиций. Сосновые леса распространены на горах, сложенных гранитами, гранитогнейсами и песчаниками, а лиственничные – на горах, сложенных глинистыми сланцами и известняками.
В долинах рек здесь широко распространены тополевые, тополево-лиственичные, яблонево-черемуховые леса, ивняки и черемушники. Нередки здесь и разнотравно-злаковые луга, встречающиеся по дну широких распадков и на относительно сырых участках надпойменных террас рек и ручьев (фото 14).

Фото 13. Внедрение сосны обыкновенной в даурскую прерию в окрестностях с. Дадал в Монголии. Фото И.Н. Сафроновой
|

Фото 14. Долина небольшой реки в окрестностях с. Дадал в лесостепном поясе
|
Нижний лесной пояс простирается до высот 1600–1700 м н.у.м. В пределах этого пояса преобладают лиственничные и лиственнично-березовые леса, реже встречаются сосновые леса, которые здесь жестко приурочены к гранитам. Даурские прерии в этом поясе встречаются изредка на крутых склонах южной и юго-восточной экспозиции и занимают небольшие участки среди леса. В долинах рек в поймах обычны тополевые и тополево-лиственничные леса, ивняки, черемушники. Для долин рек и днищ широких распадков здесь характерны заросли кустарниковой березки (Betula fruticosa, B. humilis) и кустарниковой ивы (Salix sochondinensis). Разнотравные и разнотравно-злаковые луга в этом поясе встречаются спорадически в поймах среди ивняков и тополевников.
Верхний лесной пояс в горах Хэнтея занимает высоты от 1700 до 2000 м н.у.м. Здесь господствуют кедрово-лиственичные, лиственнично-кедровые (Larix dahurica, Pinus sibirica) и кедровые леса. Изредка в долинах ручьев в составе кедрово-лиственничных и кедровых лесов встречается ель (Picea obovata) и еще реже – пихта сибирская (Abies sibirica). В поймах ручьев в местах образования зимой наледных полей в верхнем лесном поясе широко распространены ивняки и ерники, а также заросли кустарничков (Salix saxatilis, Vaccinium uliginosum).
Гольцовый, или горнотундровый, пояс в горах Хэнтея занимает высоты выше 2000–2100 м н.у.м. Здесь преобладают эпилитно-лишайниковые пустыни на каменных россыпях, а также кустарничковые (Drias oxyodontha) и разнотравно-кустарничково-лишайниковые тундры на щебнистых и щебнисто-каменистых участках. Разнотравные альпийские лужайки в гольцах Хэнтея редки и приурочены к берегам ручейков, стекающих со склонов на нагорные террасы.
В 1982–1984 гг. в Сохондинском заповеднике было заложено 30 постоянных геоботанических площадей размером 50х50 м. В закладке и первичном описании пробных площадей участвовали А.В. Галанин, А.В. Беликович, И.С. Ефимова, Н.С. Проскурина. В 2000–2005 гг. А.В. Галаниным, А.В. Беликович и И.А. Галаниной была проведена ревизия этих площадей. Таким образом, была получена информация об изменении растительности за 20 лет в гольцовом, подгольцовом, верхнем и нижнем лесных поясах заповедника. При ревизии отмечались изменения во флористическом составе растительности, обилии всех видов, изменения контуров основных синузий. Результаты ревизии показали сильные, можно сказать, катастрофические изменения в растительном покрове всех трех поясов.
В дриадово-разнотравной куртинной тундре с элементами нивальности в ложе рано стаивающего снежника появились 8 новых видов – в том числе Betula rotundifolia, Carex sedakovii, Saxifraga nelsoniana, Viola biflora, Dracocephalum grandifolium, столько же видов здесь выпало или не было найдено по причине крайне низкой их численности. Общие флористические изменения на площади довольно значительные – 16 видов из 49. В дриадово-разнотравной тундре на каменистой террасе появились 11 новых видов, многие виды сменили свое обилие. Произошла смены доминантов: тундра из полидоминантной разнотравной стала осоково-дриадово-ивковой. Так, заметно возросло обилие курильского чая, шульции, горца живородящего, ивы растопыренной, ивы барбарисолистной. Данное местообитание стало более холодным и сырым, так как на террасе в связи с потеплением интенсивнее идет оттайка многолетней мерзлоты. В куртинной разнотравно-мохово-лишайниковой каменистой тундре на вершине гольца Большое Сохондо (высота 2500 м н.у.м.) по сравнению с 1983 г., в 2000 г. исчезло 8 видов, но появились 4 новых вида.
По гари на каменистой нагорной террасе на отроге гольца Цаган-Ула идет возобновление лиственницы, однако крайне угнетенные особи лиственницы в последнее десятилетие стали усыхать. На месте редколесья формируется куртинная тундра с редкими кустарниками. Возобновившиеся к 1983 г. особи ольховника, березы плосколистной, ивы енисейской к 2000 г. полностью отмерли. Исчезли с площади кусты шиповника, куртинки голубики, 5 видов разнотравья, но появилось 18 новых видов, что говорит о сильных и необратимых изменениях экологических условий данного местообитания. В зарослях кедрового стланика на уступе нагорной террасы гольца Цаган-Ула отмечено явное усыхание кедрового стланика по всей площади. Отмерло около 20% всех кустов, зарегистрированных в 1983 г., особенно много их погибло вдоль водотоков и по окраю курумов. На этой площади за 17 лет поменялось 22 вида сосудистых растений. Изменения эти частично связаны с усыханием кедрового стланика и общей ксерофитизацией растительности, опять же из-за протайки многолетней мерзлоты.
В кустарниково-разнотравно-зеленомошном приручьевом субальпийском кедровнике на берегу Букукунского озера продолжается выпадение пихты сибирской. По всей видимости, изменился гидрологический режим данного местообитания: основной сток стал меньше, вода ушла в курум, и единый поток разделился на множество мелких потоков под камнями, в результате чего лесные ценоэлементы вдоль этих водотоков стали заменяться на элементы сырых лугов. Как следствие, мезофильные виды стали вытесняться мезогигрофитами. Видовой состав здесь изменился довольно сильно в сторону обеднения лесного разнотравья, взамен которого появились осоки.
В субальпийском кустарничково-зеленомошном с подлеском из кедрового стланика кедровнике (на второй приозерной террасе Букукунского озера) идет постепенный медленный вывал старых деревьев, в то же время также медленно идет возобновление. Мертвые стволы долгое время остаются здесь в виде сухостоя, а упав, долго лежат, не разлагаясь. Многие деревья в виде сухостоя могут находиться десятки лет. Есть сухие стволы, еще крепко стоящие, которые уже 25 лет назад были сухими. Заметна некоторая активизация на площади кедра сибирского и постепенное угасание ценопопуляции лиственницы даурской.
В кедровнике зеленомошно-бруснично-вейниковом произошла смена формулы древостоя: в 1983 г. древостой имел состав 7К3Л, а в 2000г. – 6К4Л. Здесь начинается распад древостоя, очень много стволов вывалилось и лежит на площади. Вероятно, усыхание леса – это результат иссушения склона из-за более глубокой протайки мерзлоты. Здесь активизировали свои позиции береза, брусника, багульник, княженика, малина сахалинская, вейник. Изменения в видовом составе связаны с сильным осветлением древостоя, появлением вывалов, изменением гидрологического режима.
На кедрово-лиственнично-мелколиственной разнотравно-кустарниковой редине на крутом каменистом склоне идет сильное обеднение видового состава и ксерофитизация растительности. Обеднение больше всего затронуло разнотравье (полностью выпало 11 видов), причем как на самих каменных россыпях, так и в окружающих их лесах и редколесьях. Обилие ревеня снизилось здесь с 3 до 1 балла, заметно снизили свое обилие папоротники, произрастающие в каменных нишах.
В лиственнично-кедровом баданово-бруснично-багульниковом зеленомошном лесу в вершине небольшого лога под скалами на склоне западной экспозиции 5–8 лет назад прошел небольшой низовой пожар. По-видимому, в этом пожаре выгорели все кустарники по дну лога и травянистая растительность. На площади появилось возобновление березы белой, а также рододендрон даурский, барбарис, ива копьевидная и 6 новых видов травянистых, но исчезли спирея и смородина черная (выгорели во время пожара). Судя по состоянию растительного покрова, общие условия здесь в целом остались стабильными.
Лиственнично-кедровый ерниково-бруснично-зеленомошный лес на ровной моренной поверхности за 17 лет лес превратился в ерниковый сухой лиственнично-кедровый, появилось возобновление ели высотой 1,2 м. Выпали можжевельник обыкновенный, ива растопыренная, Spiraea davurica , багульник, снизилось обилие голубики. Снижение таксономического разнообразия на площади почти в два раза свидетельствует об установлении здесь однообразных условий на всей площади, а также об общем иссушении данного местообитания. В мохово-лишайниковом покрове резко снизилось проективное покрытие зеленых мхов, на их месте доминируют кустистые эпигейные лишайники из семейства кладониевых.
Столь же существенные изменения растительности произошли и в пойменных, приручьевых местообитаниях. Например, в лиственнично-кедровом разнотравно-кустарниковом приручьевом лесу на склоне западной экспозиции появилось возобновление березы плосколистной, но резко снизили свою активность береза кустарниковая, ольха, особенно ивы (проективное покрытие ивняков уменьшилось в 2–3 раза), а также малина сахалинская. Изменения в видовом составе свидетельствуют о повышении базиса эрозии ручья и выходе большей части площади из пойменного режима в надпойменный. Основная часть стока сейчас идет по руслу, по краям же долины сток резко снизился из-за общего иссушения склона.
Общие изменения в древостое площадей верхнего лесного пояса свидетельствуют об идущей здесь смене лиственничных и лиственнично-кедровых лесов кедровыми. Кедр заметно активизировался практически на всех площадях, он преобладает в возобновлении, кроме того, в долинных лесах началась активизация ели сибирской: появилось ее возобновление, ранее не отмечавшееся. Изменения в видовом составе показывают общее иссушение региона, интенсификацию процессов оттаивания многолетнемерзлых пород и исчезновение ледяных линз в теле морен на дне Джермалтай-Ингодинской депрессии.
На остепненных лугах и в ерниках на надпойменной террасе в 2003 г., по сравнению с 1983 г., различия в растительности разных фаций стали очень малозаметны, так как ложбины высохли и заросли степными травами, потеснившими луговых мехофитов. На площади исчез 21 вид, появилось 11 новых, итого из 111 видов изменились 32 (29%). Флористические изменения отражают здесь смену гидрологического режима и постепенное снижение градиента увлажнения в зависимости от микрорельефа (комплекс ложбин/гряд). Выпало много мезогигрофитов и мезофитов, ранее произраставших в ложбинах. Появилось несколько новых видов злаков, а также ряд видов, свойственных прериям Южного Забайкалья. Сильно активизировались здесь мятлик степной и кобрезия, которые стали содоминировать во всех фациях.
Процесс остепнения затронул все фации, однако есть ряд изменений, трудно объяснимых с точки зрения ксерофитизации местообитания. Так, с площади исчез тимьян, сильно снизил свое обилие на всех фациях Androsace incana , характерный для каменисто-щебнистых субстратов. Эти и другие факты свидетельствуют о постепенном исчезновении на площади каменистых поверхностей и сильном задернении некогда обнаженных галечных субстратов. При этом луговые и степные травы начинают распределяться по площади более равномерно, исчезают различия в обилии видов между фациями.
В приручьевом ернике большая часть площади теперь не заболочена, проточное увлажнение наблюдается лишь в полосе 10 м вдоль русла ручья. Ивняки из ивы Коха по границе прирусловых зарослей в значительной степени усохли. На площади исчез 21 вид, 11 видов появилось новых. Итого поменялось 32 вида из общего числа 85. Исчезли, или сильно снизили свое обилие виды-индикаторы сильного переувлажнения застойного характера, но большинство видов разнотравья, вероятно, выпало в результате пожара. Пожар, а возможно, и понижение базиса эрозии привели к иссушению значительной части площади.
Большие изменения произошли и в самом, казалось бы, устойчивом элементе растительности лесостепного ландшафта – разнотравной горной прерии на крутом склоне. Здесь 20 лет назад основную часть площади занимал участок остепненного луга (прерии), сверху спускалась кустарниковая степь с ерниками, и по краю шла поросль осины высотой до 1–1,5 м. В настоящее время осинник разросся и занимает почти треть площади, деревья выросли до высоты 5–6 м. Ерники также получили большое развитие, сама степь стала носить луговой характер, так как исчез ряд чисто степных видов и получили распространение луговые мезофилы. Произошли изменения на уровне фациальной структуры: ранее по характеру растительности здесь были выделены три фации: кустарниковая степь, низкотравная степь, высокотравный остепненный луг. В 2001 г. выделены также три фации: осинник, ерник, остепненный луг. Число выпавших видов на этой площади – 11, число новых видов – 13. Итого 24 (25%) видов здесь поменялось. Заметно, что на остепненных участках снизилось видовое богатство, вероятно, в связи с большей сухостью климата. Сукцессия здесь явно направлена на восстановление осинового леса.
Изменения, которые мы пронаблюдали на геоботанических пробных площадях, произошедшие за последние 20 лет, поражают своим размахом и всеохватностью. Существенные, а зачастую резкие флористические изменения произошли не только на участках, явно вовлеченных в сукцессионный процесс, но и на площадях, растительность которых считается коренной, а иногда и климаксовой – в тундрах гольцового пояса, кедрово-лиственничных лесах верхнего лесного пояса, лиственничных и сосновых лесах и в лугостепях нижнего лесного пояса. В среднем флористический состав каждой площади за этот период сменился на 20–30%, а есть площади, где он изменился на 45–55%. На многих площадях достоверно изменилось обилие видов, в том числе и видов доминантов и эдификаторов. Есть основания полагать, что эти изменения связаны с резкой аридизацией климата Центральной Азии в конце ХХ столетия.
Сильно выгорели и усохли в долинах рек заросли кустарниковых берез – ерники, а на их месте появились роскошные луга с преобладанием копеечника и астрагала (фото 15). В субальпийском поясе сильно усох кедровый стланик, а в тундровые сообщества проникли виды трав из лесостепного пояса. На каменных россыпях в лесном и субальпийском поясах заметно активизировался барбарис сибирский. Даже на вершине гольца Сохондо растительность заметно изменилась.

Фото 15. Долина реки Онон близ российско-монгольской границы. Лесостепной пояс. Фото Л.М. Долгалевой |

Фото 16. Березовый лес из березы плосколистной на месте даурской прерии. Монголия. Фото Л.М. Долгалевой |
Граница между лесом и прерией в пространстве ландшафта Южного Забайкалья постоянно сдвигается в зависимости от многолетних климатических циклов (фото 11). В настоящее время в связи с циклом иссушения климата все большее число лиственничных лесов горит, и поэтому активизируется степной компонент растительности, а лесной деградирует. В периоды с влажным гумидным климатом прерия сменяется лесом, лес в это время наступает на прерию. Результаты ревизии растительности постоянных пробных площадей в заповеднике однозначно говорят о ксерофитизации растительности Южного Забайкалья и Северной Монголии в последние десятилетия. По наблюдениям Никиты Петровича Соколова на гольце Сохондо в середине ХVIII столетия (250 лет назад), климат был более гумидным, в результате чего верхняя граница леса тогда проходила здесь на 150 м выше, чем в настоящее время.
В процессе заселения даурской прерии лесом (фото 16) из состава травяного покрова выпадают степные ксерофиты: ковыль Крылова, овсяница ленская, володушка, мордовник даурский, житняк гребенчатый и другие виды. Появляются же лугово-лесные и лугостепные растения мезофиты и ксеромезофиты: василистник малый, овсяница овечья, клевер люпинолистный, лапчатка пижмолистная, мятлик сибирский, а для более сомкнутых лесов характерны осока ланцетовидная, кровохлебка лекарственная, горец узколистный, дендрантема Завадского, горец лисохвостниковый, сон-трава Турчанинова, земляника восточная, герань луговая, колокольчик скученный, копеечник, аконит бородатый и др. Если на степных участках доминируют ковыль тырса и другие степные злаки и разнотравье, то в сомкнутых насаждениях доминирование переходит к осоке ланцетной и лугово-лесному разнотравью. При смене леса прерией, и, наоборот, при смене прерии лесом изменяется, естественно, и почвообразовательный процесс. К сожалению, эти изменения почвообразовательного процесса в Южном Забайкалье и в Северной Монголии почвоведами совсем не изучались.
Полученные выводы подтверждаются и лихенологическими исследованиями в субальпийском и лесостепном поясах Сохондинского биосферного заповедника. Изучение размерной структуры популяций лишайников эпифитов в 2001 г. позволяют говорить о наличии в этом регионе также 600–700-летнего климатического цикла, в течение которого гумидная эпоха, в которую на горе Сохондо был Никита Соколов, сменилась в настоящее время аридной. Примерно через 300 лет здесь должна произойти снова гумидизация климата и активизация лесной растительности. Максимум гумидности имел место в ХI и ХII вв., а вот в ХIII в. здесь началось иссушение климата. Не оно ли толкнуло монголов тогда искать пастбища для скота и теснить своих восточных и западных соседей? Еще раньше, и тоже на 600–700 лет, иссушение климата вынудило переселяться на запад тюрков, а до этого, и тоже минус 600–700 лет, была мощная волна миграции из Центральной Азии народа, известного китайцам как сю-ну, а европейцам как гунны.
В июле 2001 г. нам удалось обследовать накипные лишайники на каменных плитах на древних захоронениях в окрестности станицы Кыра. Мы измерили поперечники всех особей на десятках каменных плит и установили, что популяции лишайников эпифитов на могильных плитах в течение последних двух тысячелетий пополнялись неравномерно, по-видимому, в соответствии с тем же климатическим циклом. Деформации кривых частотного распределения особей накипных лишайников в зависимости от их размерной характеристики (максимального поперечника особи) оказались достоверными при вероятности ошибки Р=0,05. Очень похоже, что те особи лишайников, которые поселились на камнях после переноса этих камней на могилу, смогли вырасти до максимального размера с поперечником 5–6 см. Если учесть, что поперечник здешних накипных лишайников эпилитов в среднем увеличивался на 3 мм за каждые 100 лет (именно такую оценку скорости роста накипных эпилитов дают специалисты-лихенологи), то следует признать, что возраст плиточных могил в Южном Забайкалье не менее 2000 лет. Разумеется, это очень приблизительная оценка, требующая дальнейших уточнений и проверок. Но это первая оценка возраста плиточных захоронений, основанная хотя бы на каких-то объективных данных.
Литература
• Беликович А.В. Растительный покров северной части Корякского нагорья. – Владивосток: Дальнаука, 2001. – 420 с.
• Беликович А.В., А.В. Галанин. Забайкальский харганат как тип растительности // Комаровские чтения. – Владивосток: Дальнаука, 2005. – Вып. 39.– С. 162–185.
• Быков Б.А. Введение в фитоценологию. – Алма-Ата: Наука, 1970. – 234 с.
• Воронов А.Г. Геоботаника – М.: Наука, 1973. – 213 с.
• Высотная поясность // Википедия. Адрес доступа: http://ru.wikipedia.org /wiki/
• Беликович А.В., Галанин А.В. Изменения в растительном покрове Сохондинского заповедника по результатам ревизии геоботанических пробных площадей (1983-2001) // Растительный и животный мир Сохондинского государственного заповедника: труды Сохондинского биосферного заповедника. Вып.1. – Чита: изд-во СБЗ, 2002. – С.14–34.
• Галанин А.В. 1989. Ценотическая организация растительного покрова. – Владивосток: Изд-во ДВО РАН, 1989. – 167 с.
• Галанин А.В. Флора и ландшафтно-экологическая структура растительного покрова. – Владивосток: ДВО АН СССР, 1991. – 272 с.
• Галанин А.В. Ценотическая организация растительного покрова. – Владивосток: БСИ ДВО РАН, 2005. (Электронное издание монографии на сайте БСИ ДВО РАН) (по адресу: gardens.narod.ru).
• Дулепова Б.И. Петрофильные кустарниковые степи, их значение и охрана // Охрана природы и воспроизводство естественных ресурсов. – Чита, 1979. – С. 95–97.
• Камелин Р.В. Флороценотипы растительности Монгольской Народной Республики // Ботан. журн., 1987. Т. 72. № 12. – С.1580–1594.
• Куминова А.В.Растительный покров Алтая. – Новосибирск: СО АН СССР, 1960. – 450 с.
• Маркевич В.С. Палиностратиграфия меловых отложений Северо-Востока СССР // Континентальный мел СССР. – Владивосток: ДВО РАН, 1990. – С. 132–143.
• Огуреева Г.Н. Ботаническая география Алтая. – М.: Наука, 1980. – 189 с.
• Пармузин Ю.П. Тундролесье СССР. – М., Мысль, 1979. – 295 с.
• Природные зоны и высотная поясность // Сайт Экологический центр «Экосистема». Адрес доступа: http://www.ecosystema.ru/ 08nature/world /geoussr/31-1.htm.
• Сочава В.Б. Онон-Аргунская степь как объект стационарных физико-географических исследований // Алкучанский Говин. – М.; Л.: Наука, 1964. – С. 3–23.
• Сохондинский биосферный заповедник // Сайт «Культурный мир». Адрес доступа: http://www.kulttur.com/index.
• Тюлина Л.Н. Влажный прибайкальский тип поясности растительности. – Новосибирск: Наука, 1976. – 319 с.
• Шенников А.П. Введение в геоботанику. – Л.: Изд-во ЛГУ, 1964. – 448 с.