УДК 58.0558.02
ДИНАМИЧЕСКАЯ ПАРАДИГМА В ГЕОБОТАНИКЕ И БОТАНИЧЕСКОЙ ГЕОГРАФИИ
А.В. Галанин, И.А. Галанина
A.V. Galanin , I.A. Galanina. DYNAMIC PARADIGM IN GEOBOTANY AND BOTANICAL GEOGRAPHY. The article presents the advantages of dynamic paradigm in vegetation studies in plant ecology and botanical geography. The authors review the history and dynamics of flora and vegetation of the Russian Far East associated with cyclical changes in climate in Cenozoic and reduction of the Tetis Sea.
Tabl. 2. Bibl. 20.
Растительный покров любой территории – это функция четырех основных факторов: 1 – предшествующей его динамики (эффект прошлого), 2 – современного климата, 3 – современного рельефа, 4 – геологической динамики местности. Большинство современных геоботаников растительный покров рассматривают как объект, существующий в пространстве, в то время как он является процессом, и вне времени понять закономерности многообразия растительного покрова невозможно. Растительный покров как процесс, реализующийся в пространстве и времени, – это суть динамической парадигмы в геоботанике и ботанической географии. Нам могут возразить, что же тут нового? Ведь флорогенез, процессы видообразования, миграции видов и флор изучали испокон веков. Да, это так, динамику флор, видов, сообществ изучали еще в XVIII веке, но когда начинали классифицировать и картировать растительные сообщества и их пространственные комбинации, об этом, как правило, забывали. Разграничить растительные сообщества в пространстве намного проще, чем разграничить их во времени.
Массовое увлечение геоботаников флористическими методами классификации растительности настолько формализовало разграничение растительных сообществ, что сделало такую классификацию непригодной не только для изучения динамики растительного покрова, но и для его картирования. Е.Н. Лукашова (статья в БСЭ) писала: «Географическая закономерность дифференциации ландшафтной оболочки Земли, проявляющаяся
в последовательной и определенной смене географических поясов и зон, обусловленной, в первую очередь, изменениями количества лучистой энергии Солнца, падающей на поверхность Земли, в зависимости от географической широты. Зональность присуща и большинству компонентов и процессов природных территориальных комплексов – климатическим, гидрологическим, геохимическим и геоморфологическим процессам, почвенному и растительному покровам и животному миру, отчасти образованию осадочных пород. Уменьшение угла падения солнечных лучей от экватора к полюсам вызывает выделение широтных радиационных поясов – жаркого, двух умеренных и двух холодных. Формирование аналогичных тепловых и тем более климатических и географических поясов связано уже со свойствами и циркуляцией атмосферы, на которые большое влияние оказывает распределение суши и океанов (причины последнего азональны). Каждому географическому поясу присущ свой набор (спектр) зон и их последовательность». Зональность тепловых условий установил еще в начале XIX века Александр Гумбольдт (1962), а в качестве основного закона дифференциации крупных природных комплексов зональность определена в 1898 г. В.В. Докучаевым.
Между растительным покровом и климатом прослеживается отчетливая связь, которая нашла отражение в понятии биоклиматической зональности (Григорьев, Будыко, 1956; Григорьев, 1957). Говоря о климате, мы имеем в виду два главных системы образующих фактора – количество и режим получения местностью тепла и количество и режим получения ею осадков. Гидротермическая система весьма сложна, поэтому обойтись выделением широтных зон растительности ботаники не могут. Схема природных зон, учитывающая только годовые суммы температур и суммы осадков, недостаточна настолько, что ее использование зачастую теряет всякий смысл. При одинаковости указанных параметров растительный покров бывает совсем не одинаков. Например, широколиственные леса в окрестностях г. Осло в Скандинавии, дремучие еловые леса в Вологодской области и лиственично-сосновые леса в южной Якутии. Однако теория настолько сильно довлеет над нами, что мы этим фактам не придаем большого значения. В пределах зоны бореальных лесов мы рассматриваем и темнохвойную тайгу северной Европы, и светлохвойную тайгу восточной Сибири, которые между собой по характеру биологического круговорота вещества совершенно различны. Или другой пример: степи Европы и степи Центральной Азии обычно относят к одной зоне степей. Но в европейских степях самая большая биогеохимическая активность имеет место весной и в начале лета, а в центральноазиатских – во второй половине лета и начале осени.
Другой биоклиматический вопрос можно сформулировать так: что важнее для растительного покрова, фактор «тепло», или фактор «увлажнение». Большинство геоботаников и ботаникогеографов отдают предпочтение фактору «тепло», а фактор «увлажнение» ставят на второе место. Од нако линейной зависимости между растительным покровом и этими факторами нет. Более важен для растительности тот фактор, который находится в минимуме. Много тепла, но мало влаги – растительность скудная в виде сухих степей. Много влаги, но мало тепла – растительность тоже скудная в виде полярных тундр. Крайности сходятся, не случайно в горах Хэнтея участки степной растительности проникают в пояс горных тундр, но отсутствуют в поясе хвойных лиственничных и кедровых лесов.
Но не только современный климат определяет характер растительности. Растительность своим видовым составом, составом сообществ отражает и прошлое состояние растительного покрова. Например, современные еловые леса в хребте Сихотэ-Алинь сформировались в условиях климата XVIII–XIX веков. В конце XX века климат изменился, и еловые леса стали «неадэкватны» новым климатическим условиям. Ель стала усыхать. В новых условиях на этих элементах горного ландшафта стали оптимальны леса из кедра корейского, но еловые леса все еще удерживают позиции. Потребуется не менее 100 лет, прежде чем произойдет их деградация, и место ели аянской займет кедр корейский. При этом спутники ели аянской в сообществах с ее доминированием перейдут в сообщество с доминированием кедра корейского и несколько столетий будут удерживать позиции в новом для них сообществе. Для растительного покрова и конкретных растительных сообществ присуща немалая ценотическая инерция.
В том случае если климат изменяется циклически с длительностью циклов 300–700 лет, то сукцессионный процесс не успевает закончиться, как начинается новый сукцессионный процесс. При этом виды из первого сообщества, изменив свою ценотическую активность, сохраняются и в новом сообществе. А когда прежние климатические условия вернутся, они вновь активизируются. Такой климатический ритм поддерживает высокое видовое разнообразие растительных сообществ, которое мы называем их сверхразнообразием. Таким образом климаксовая растительность в большинстве регионов Земли, где климат циклически изменяется, сформироваться не может, так как сукцессия одной фазы климатического цикла не успевает закончиться, как начинается другой. При перманентном неравновесном состоянии растительных сообществ говорить о какой-то их ценотической замкнутости просто бессмысленно.
Обычно геоботаники считают, что геологические процессы прямым фактором, влияющим на характер растительного покрова, не являются, так как эти процессы идут очень медленно. Горы воздымаются со скоростью несколько сантиметров в год, пенепленизация гор тоже происходит медленно. Однако наши наблюдения позволяют говорить обратное. Например, тектоническое опускание одного края земной плиты всего на полметра может привести к заболачиванию одной части широкой речной долины и иссушению другой. Причем изменения происходят буквально в течение десятилетий. Подобные процессы мы наблюдали в 1996 г. в северной части Корякского нагорья, где в связи с тектоническим опусканием правой части широкой долины реки Конрарывеем, ивняки первой надпойменной террасы стали заболачиваться, а такой же террасы по левому берегу иссушаться и сменяться кустарничково-лишайниковыми тундрами. Похоже, что движения земной плиты, подстилающей рыхлые отложения долины реки, периодически изменяют направление своего движения: то правый борт долины воздымается, а левый борт опускается, то левый поднимается, а правый опускается. Предполагать такой тектонический ритм позволяют некоторые виды растений гигрофитов (Caltha arctica, Carex aquatilis), которые ухитряются выживать на террасе по левому борту долины и виды растений мезоксерофитов (Salix xerophylla, S. glauca), которые встречаются в заболоченной правой части долины. При таких тектонических колебательных движениях русло реки сдвигается то к одному борту долины, то к другому.
С тектоническими сжатиями и растяжениями земной коры могут быть связаны колебания уровня многих озер и грунтовых вод. Например, с 30-летним циклом изменяется уровень Торейских озер в Даурии. При этом достоверной сколь-либо значимой зависимости обводненности озер от количества выпадающих осадков не наблюдается. При сжатии земной коры содержащаяся в ней вода буквально выжимается вверх, при этом довольно резко изменяется уровень грунтовых вод, что в свою очередь отражается в изменении растительности. Давно замечено, что перед землетрясениями резко изменяется уровень воды в скважинах и колодцах. Так, в Кыринской котловине за последние 20 лет уровень грунтовых вод снизился на 10 м, при этом высохли колодцы, а продуктивность пастбищ снизилась в разы. Объяснить эти процессы только уменьшением количества выпадающих осадков невозможно, их количество снизилось только на 15–20%.
Современная флора и растительность большинства территорий формировалась и существует в режиме циклических изменений, связанных не только с циклическими изменениями климата, но и с циклическими микротектоническими движениями земной коры. Поэтому в конкретной флоре можно выделить несколько генетических элементов – групп видов, экологически оптимальных для разных этапов климатических и тектонических циклов. Если учесть, что циклы с малым периодом накладываются на циклы с большим периодом, в результате чего образуется сложная гармоника, которую бывает сложно разделить на элементарные составляющие, то разобраться в генетической структуре флор бывает не так просто. Но ясно одно, чем более подвержен ритмическим изменениям климат местности, тем сложнее генетическая структура его флоры, и чем стабильнее климат, тем проще эта структура, тем ценотически закрытее конкретные растительные сообщества.
Надо заметить, что разные растительные сообщества обладают разной степенью ценотической закрытости. Например, видовой состав растительных сообществ верховых болот на севере Восточной Европы изменился меньше, чем видовой состав бореальных лесов. На болотах как росла чахлая жизненная форма сосны обыкновенной (Pinus sylvestris), так и растет до сих пор. С конца последнего ледникового периода на болотах растут карликовая березка (Betula nana), голубика (Vaccinium uliginosum), клюква (Oxicoccus palustris), морошка (Rubus chamaemorus) и др. виды. В лесах же за этот промежуток времени береза (Betula pubescens) сменилась елью сибирской (Picea obovata), ель – сосной (Pinus sylvestris), сосна снова елью, но елью европейской (Picea abies), в ряде районов в хвойные леса внедрились липа сердцелистная (Tilia cordata), клен остролистный (Acer platanoides), дуб черешчатый (Quercus robur). Изучая конкретные флоры чукотской тундры, мы обнаружили, что растительность склонов гор ценотически менее закрыта, чем растительность тундровых болот и равнинных кочкарных тундр с доминированием пушицы влагалищной (Eriophorum vaginatum) и осоки мечелисной (Carex ensifolia ssp. lugens).
В масштабах геологического времени растительный покров соседних территорий то конвергировал по климатическим условиям и при этом сближался по составу и строению, то дивергировал (расходился). В момент конвергирования (схождения) происходил обмен видами и флористическими комплексами (ассоциациями видов), при этом флоры территорий становились более сходными вплоть до их полного объединения. При дивергировании климата флоры и растительность территорий становились все более различными, так как на разошедшиеся по климату территории приходили разные мигранты, а популяции общих видов эволюировали в разных направлениях, становясь разными подвидами и викарирующими видами. Быстрее всего при климатической дивергенции изменялся состав видов доминантов, а малоактивные виды сохранялись, лишь слегка снизив, или увеличив свою активность. Например, современная растительность Даурии, если судить по доминирующим видам, существенно отличается от таковой Манчжурии, но если судить по сходству флор в целом, то растительный покров и Даурии, и Манчжурии принадлежат к одной Восточноазиатской ботанико-географической области. Это показано нами при сравнительном анализе ряда семейств из флор Даурии и Манчжурии (Галанин, 2006; Галанин и др., 2009).
Геоботанические области обычно делят на провинции, а последние на округа и округа на районы. Сколь-либо четких критериев отнесения подразделения к провинции, округу или району не существует. На практике разные авторы при ботаническом делении территории используют различные критерии, причем об этих критериях ничего вразумительного не сообщают, отчего схемы районирования, построенные разными авторами, получаются несопоставимыми. В результате положение границ выделов у разных авторов может совпадать на 70-80%, но ранг выделов при этом не совпадает никогда.
Понятие «тип растительности» в геоботанике должно занимать центральное место. Главными характерными особенностями типа растительности должны быть объем, скорость и степень замкнутости биологического круговорота, обеспечиваемого растительными сообществами этого типа в экологических системах. Эти параметры биологического круговорота вещества в наибольшей степени зависят от характера жизненных форм растений, сообитающих в конкретных растительных сообществах (Галанин, 1989, 1991). Поэтому, характеризуя конкретное растительное сообщество или участок растительного покрова в ранге фитоценоза, геоботаник обязан назвать жизненную форму доминирующих растений. Известно, что луга – это сообщества с доминированием многолетних травянистых мезофитов, степи – с доминированием травянистых ксерофитов: плотнодерновинных злаков, осок и луковичных и корневищных эфемероидов, светлохвойные леса – листопадных хвойных деревьев, темнохвойные – вечнозеленых хвойных деревьев, неморальные широколиственные леса – листопадных широколиственных деревьев. Каждый тип растительности имеет свою историю, насчитывающую десятки, а иногда и сотни миллионов лет, в течение которых происходила коэволюция совместно обитающих видов растений и животных. Эта эволюция корректировалась определенными модусами эвлюции – определенными климатическими и экологическими условиями внешней среды, с которыми данный тип растительности находился в относительном биогеохимическом равновесии. В результате для каждого типа ландшафта сформировался свой особый тип экосистем и свой особый тип растительности.
В умеренной зоне при дефиците воды в течение почти всего вегетационного периода, но при относительно хорошем увлажнении весной за счет тающего снега и дождей устойчивым типом растительности являются степи, которые расцветают за счет эфемероидов весной, но летом сильно усыхают. В степях запас фитомассы над поверхностью почвы в 7–8 раз меньше, чем в почве (Сочава, 1986). В умеренной зоне при отсутствии устойчивого дефицита воды в течение вегетационного периода там, где выражен аллювиальный процесс, устойчивым типом растительности являются луга. На лугах запас подземной фитомассы только в 2–3 раза больше, чем надземной. Для типа растительности верховое болото характерен чрезвычайно замедленный и несбалансированный биологический круговорот, в результате чего в болотной экосистеме происходит накопление торфа. В темнохвойной тайге значительная часть фитомассы накапливается в стволах и сучьях, и на многие десятки лет вещество выводится из ежегодного биологического круговорота. Хвоя в таких лесах тоже функционирует 3–4 (5) лет, что также приводит к существенному замедлению биологического круговорота. При большом запасе фитомассы темнохвойная тайга, в отличие от луга, обеспечивает замедленный круговорот вещества. По скорости круговорота темнохвойные леса ближе к сухим степям и полупустыням, но по типу «складирования органического вещества "надолго" они от них существенно отличаются.
Темнохвойная тайга устойчива в условиях бореального климата с относительно теплым летом, холодной зимой и достаточным в течение вегетационного периода увлажнением. В светлохвойной тайге хвоя отмирает ежегодно, поэтому биологический круговорот здесь более интенсивный, чем в темнохвойной тайге. Интенсификации круговорота вещества в светлохвойной тайге способствует то, что весной, пока хвоя на деревьях не отросла полностью, под осветленным пологом леса интенсивно развиваются кустарники и травы, которые весьма интенсивно участвуют в обеспечении ежегодного биологического круговорота. Светлохвойная тайга устойчива в условиях резко континентального климата с очень холодной зимой и теплым летом, где зимой выпадает мало снега, почва сильно промерзает так, что нередко образуется вечная мерзлота, а весна и начало лета отличаются сухостью.
Условимся под климатом понимать устойчивый, усредненный за 40–50 лет, годовой режим погоды (режим температуры, осадков, ветров, давления атмосферы), который связан с обращением Земли вокруг Солнца, наклоном земной оси к плоскости эклиптики, географической широтой и географической долготой местности. В этом случае суточный цикл погоды – это суточный режим температуры, осадков, ветров, давления атмосферы. Ну а погода – это одномоментная характеристика перечисленных параметров в некоторой местности. Действительно, погода может измениться в течение часа. Один суточный цикл погоды бывает похож, а бывает и не очень, на такой же цикл через сутки и более.
В Восточной Сибири, на Дальнем Востоке и в Центральной Азии в аридные фазы климатических циклов активизировался криофильно-степной комплекс видов, и начиналась миграция видов из Центральной Азии в Якутию, на Чукотку и через Берингийский мост суши в Аляску и далее в Америку, а также на восток в Манчжурию на Сихотэ-Алинь и в Корею. В гумидные фазы активизировался другой вектор миграции видов – с Дальнего Востока на запад в Манчжурию и Даурию, из Южной и Западной Сибири в Прибайкалье и в Даурию. Поэтому Восточноазиатская ботанико-географическая область вытянута вдоль гумидного вектора и обязана своим происхождением и существованием тихоокеанскому муссону, то затухающему, то активизирующемуся. Вдоль аридного вектора также сформировался определенный хорион растительного покрова от пустыни Гоби до пустынь запада Северной Америки (пустыня Большого Бассейна, пустыня Мохаве).
Кроме векового (600–700-летнего) климатического цикла, существует и более длительный 40700-летний, связанный с раскачиванием земной оси относительно плоскости эклиптики (плоскости в которой находится орбита Земли). Когда наклон оси к плоскости эклиптики уменьшается, то
сокращаются теплые и холодные пояса, но зато расширяются два пояса умеренного климата. Похоже, что чередование ледниковых и межледниковых эпох в плейстоцене происходило именно с такой периодичностью (Галанин, 2001).
С периодичностью 92000 лет орбита Земли то вытягивается, то снова приближается к круговой. Когда эксцентриситет орбиты увеличивается, то летние месяцы в северном полушарии сильно отличаются от летних меся-цев в южном полушарии по количеству получаемой солнечной энергии. Когда эксцентриситет уменьшается, то количество солнечного тепла, получаемое северным и южным полушариями выравнивается. Цикличность во Вселенной – движитель ее эволюции, благодаря цикличности время является не скалярной, а векторной величиной. Колебания климата на Земле в направлении «гумидность-аридность» вызывали миграции видов то в сторону материка, то в сторону океана. Изменения климата в направлении «похолодание-потепление» приводили к миграциям видов с севера на юг и обратно. В соответствии с этими изменениями климата изменялся и вектор естественного отбора. Мигрировали не только отдельные виды, но целые эколого-ценотические комплексы видов, приспособленных к совместному существованию – такие, как комплекс видов светлохвойной тайги, гипо-арктический комплекс, комплекс степных видов и т.д. Проводниками северных видов и комплексов на юг служили горные цепи, а южных на север – долины рек и южные склоны гор. Если выявить ареалогическую структуру локальных флор и их эколого-ценотическую структуру и провести их сопряженный анализ, то можно увидеть в структурах флор следы прошлых климатических эпох.
При изменениях климата одни виды растений оказывались более подвижными, чем другие, они мигрировали быстрее. Такие виды имеют специальные приспособления к переносу семян ветром на большие расстояния (виды родов Salix, Populus, Epilobium, Taraxacum и др.). Быстро распространяются виды растений, имеющие приспособления для переноса семян и плодов животными либо в виде прицепок за их шерсть, либо к перенесению в желудках. Медленнее других распространяются такие виды, как кедр корейский (Pinus koraensia), кедр сибирский (P. sibirica), кедровый стланик (P. pumila), орех маньчжурский (Juglans mandshurica). К сожалению, при анализах флор ботаники редко обращают внимание на скорости расселения видов и имеющиеся у них приспособления для этого. Без учета скорости расселения мы не можем утверждать, что всякий вид с большим ареалом старше вида с меньшим ареалом. Для расселения видов, семена которых распространяются ветром, важно в какую сторону дуют ветры в период созревания семян. Виды растений, семена которых распространяются в желудках перелетных птиц, с большей скоростью будут распространяться на юг, чем на север, так как перелетные птицы осенью летят на юг.
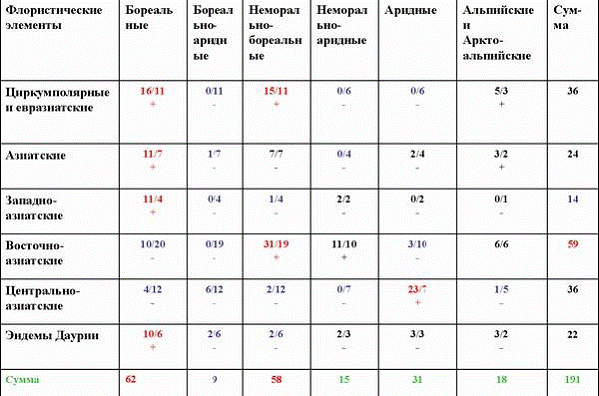
Таблица 1
Сопряженный анализ долготной и широтной ареалогической структуры дендрофлоры Даурии
|
В качестве примера рассмотрим результаты сопряженного анализа структуры дендрофлоры Даурской подобласти Восточноазиатской ботанико-географической области. Западная граница этой области совпадает с западной границей подобласти и выражена довольно резким флористическим градиентом, проходящим по восточному макросклону хр. Хамар-Дабан и западному макросклону Хангая. Восточная граница Даурской подобласти проходит по западному макросклону Большого Хингана. Южная граница совпадает с северной границей пустыни Гоби, а северная граница подобласти совпадает с северной границей области и проходит по северной границе распространения даурских степей прерий, приуроченных к южным склонам гор в Забайкалье.
Многие виды с восточноазиатским типом ареала выклиниваются на долготе Байкала, а западноазиатские виды имеют восточный предел своих ареалов также вблизи этой долготы (Беликович, Галанин, 2005; Галанин и др., 2005; Наумов, 2009). Из табл. 1 видно, что циркумполярные виды достоверно положительно сопряжены с бореальными и неморально-бореальными, азиатские и западно-азиатские – только с бореальными, восточноазиатские – с неморально-бореальными, центральноазиатские – с аридными, а эндемы Даурии – с бореальными. Неморально-аридные, альпийские и аркто-альпийские виды ни с какой долготной ареалогической группой не сопряжены. Это позволило нам утверждать, что дендрофлора Даурии в своем генезисе теснее была связана с Восточной Азией, чем с Западной, что миграции видов с востока активизировались в теплые фазы гумидных климатических эпох. |
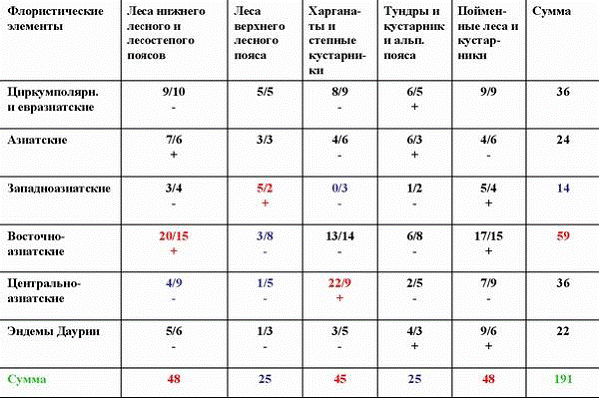
Таблица 2
Сопряженный анализ долготной ареалогической и эколого-ценотической структуры дендрофлоры Даурии
|
А вот в холодные фазы гумидных климатических эпох в Даурию мигрировали виды с запада и севера. В аридные фазы шло распространение на север центральноазиатских видов. В целом же в дендрофлоре преобладают виды с восточноазиатскими ареалами, что и побуждает рассматривать Даурию как подобласть Восточноазиатской области.
Из табл. 2 видно, что между долготной ареалогической структурой дендрофлоры Даурии и ее эколого-ценотической структурой также существует сопряженность. Так положительно сопряжены восточноазиатские виды с лесами нижнего лесного и лесостепного поясов. А вот с лесами верхнего лесного пояса положительно сопряжены западноазиатские виды. Это говорит о том, что растительность верхнего и нижнего лесных поясов в Даурии развивалась по-разному. Нижний лесной пояс здесь генетически «тяготел» к Восточной Азии, а верхний лесной – к Западной Азии. Харганаты и степные кустарники положительно сопряжены только с центральноазиатскими видами, а вот высокогорные тундры и пойменный комплекс не имеют достоверных сопряженностей не с одним из ареалогических элементов флоры. Этот анализ показывает структуру миграционных потоков видов и их приуроченность к разным эколого-ценотическим комплексам и к разным фазам климатических циклов (Галанин и др., 2005). |
Когда-то обширные по площади внутренние водоемы со значительными глубинами на юге Монголии были связаны с водоемами северного Китая и охватывали значительную территорию в центре Азии. Их исчезновение связывают с постепенным воздыманием всей территории Центральной Азии в кайнозое, приведшим к возникновению пустыни Гоби. Гоби поднята над уровнем моря на высоту от 800 до 1700 м, а хребты Гобийского Алтая поднимаются на высоту 3000 м н.у.м. Гоби является самым резко континентальным местом на планете. Температура воздуха в июле здесь поднимается до +50°, а зимой опускается до -40°. До поднятия Гималаев Центральная Азия была равниной, невысоко воздымавшейся над уровнем моря, открытой теплым влажным ветрам с океана Тетис, которые тогда проникали далеко на север. В то время территория нынешней Центральной Азии была покрыта субтропическими лесами.
Затем произошло катастрофическое сжатие Земного шара, океан Тетис начал схлопываться, а океаническая кора на его дне начала сминаться в складки и разламываться. В итоге на месте океана образовались высокие горные цепи – Гималаи, Кунь-Лунь и др. Этими горами южный суперматерик Гондвана оказался прочно припаянным к Лавразии. В эпоху сжатия впадины океанов и морей уменьшились, и моря затопили многие низменности и равнины, высота которых была менее 100–150 м.н.у.м. При последующем расширении Земного шара, зона спрединга образовалась не на месте Гималаев, а южнее. Боль-шие куски Гондваны в виде полуостровов Индостан и Индокитай остались «припаянными к Лавразии и стали ее частями, а новый окена – Индийский – возник много южнее прежнего Тетиса.
Расширение Земли привело к образованию новых глубинных разломов и зон спрединга, но там, где сиалическая материковая кора была особенно прочной, возникли не зоны спрединга, а сводовые куполообразные поднятия вроде Тибета и горных систем на территории Монголии и южной Сибири. В режиме растяжения земной коры по глубинным разломам на поверхность вытекала раскаленная и жидкая базальтовая лава. Такие потухшие щитовые вулканы и базальтовые покровы довольно часто встречаются в разных районах Центральной Азии, в том числе и в Монголии. Гималаи изолировали Центральную Азию от затока теплого влажного воздуха из акватории Индийского океана. По мере активизации горообразования происходила аридизация климата, а затем началось и опустынивание. Воздымание Тибета привело к резкому похолоданию на его поверхности и в его окрестностях. При этом, естественно, изменился и вектор эволюции растений и животных, активизировались миграции засухоустойчивых видов их аридных районов Средиземья и холодоустойчивых видов из Беринги.
Плейстоценовое оледенение в Азии было значительно меньшим по сравнению с Европой и Северной Америкой, хотя масштаб похолодания здесь был большим. Аридный климат препятствовал возникновению ледников. Они формировались только на высоких горах, а равнины и невысо кие горы были свободны от ледников. Но здесь развивалось подземное оледенение – вечная мерзлота. Мощность слоя вечной мерзлоты достигает нескольких сотен метров.
В Кайнозое сформировалось Древнесредиземноморское флористическое подцарство Голарктического флористического царства. Флора и растительность этого подцарства вначале были тропическими и были распространены по северному побережью океана Тетис. Но по мере его схлопывания происходила аридизация и формирование ксерофильной растительности в виде субтропической саванны на западе и лесостепи на востоке. Это флористическое подцарство протянулось от Макаронезии (Канарских островов в Атлантическом океане) на западе через все Средиземноморье, Переднюю и Среднюю Азию до Монголии и Китая на востоке. Еще А. Энглер в 1881 г. ввел понятие "Флора древнего Средиземноморья" (Тахтаджян, 1978). Впоследствии М.Г. Попов (1929) более детально развил идею о единой древнесредиземноморской флоре, расширив границы ее влияния вплоть до Калифорнии и Мексики.
Территория северного побережья высыхающего океана Тетис в кайнозое была ареной переселений и смешения тропических и внетропических флор, но большинство видов растений здесь имеет голарктическое происхождение. Смятое в складки дно Тетиса изолировало флору северного побережья Тетиса от флоры его южного побережья. Это особенно хорошо видно при сравнении флор Индии и Тибета. На западе схлопывающегося океана Тетис изоляция северных лавразийских флор от флор гондванских была более слабой, поэтому во флорах Макаронезийской и Средиземноморской областей довольно много африканскх (гондванских) родов и видов. Во флорах Ирано-Туранской и Центральноазиатской областей индийских (гондванских) родов и видов очень мало. Во флоре Центральноазиатской флористической области насчитывается едва ли более 5000 видов, однако, эндемизм на уровне видов и на уровне родов здесь весьма большой.
Своеобразный аридный центр видообразования в палеоген-неогене сложился в области сочленения Ангарского и Берингийского горных узлов. Этот регион оказался защищенным от воздействия влажных воздушных масс как с запада – с Атлантики, так и с востока – с Тихого океана. Большую роль как центры видообразования играли территории Даурии, Якутии, Берингии. Из Беринги, по мнению М.Г. Попова (1963), расселялись на юг и запад виды родов Euphrasia, Atraphaxis, Ceratoides, Echinops, Goniolimon, Oxytropis, Achnatherum. В плиоцене, по всей вероятности, существовал весьма протяженный транзитный (через Берингию) пояс пустынно-степной флоры от Гоби до пустынь Северной Америки. Но в Плейстоцене, по мере похолодания климата в даурско-якутско-берингийской части Великой Аридной Дуги, векторы эволюции на юге и севере этой дуги и в Азии и в Америке перестали совпадать – дуга стала распадаться. Но значительная ее часть все же сохранилась, но уже как дуга крио-аридной растительности, простиравшаяся от Даурии через Якутию и Берингию до северо-запада Северной Америки.
В плиоцене широколиственные ильмовые из Ulmus pumila леса и абрикосники из Armeniaca sibirica начали трансформироваться в харганаты (Галанин, Беликович, 2005, 2006). В сухих лесах в бассейне Селенги создались условия для генезиса степных эндемиков Даурии – Festuca dahurica, Vicia tsydenii, Caragana buriatica, Potentilla ozjorensis. Реликтов ледниковой плейстоценовой эпохи, объединяемых в «плейстоценовый флористический комплекс», много: Selaginella sanguinolenta, Festuca kryloviana, Carex amgunensis, Androsace incana, Potentilla kryloviana, P. sericea, Eremogone capillaries, E. formosa, Patrinia sibirica, Bupleurum multinerve и др. Перигляциальные степные реликты широко распространены в горах южной Сибири и северной Монголии (Соболевская, 1958; Намзалов, 1999). Криофитный морфогенез в плейстоцене проявился не только вблизи ледников, но в первую очередь в холодных ультраконтинентальных депрессиях Байкальского обрамления и в Якутии.
Таким образом флора Восточноазиатско-Западноамериканской аридной дуги в третичном периоде (палеогене и неогене) тесно взаимодействовавшая с средиземноморской флорой, в плиоцене и плейстоцене превратилась в самостоятельный криоаридный центр видообразования. Даурия, располагаясь на стыке атлантического и пацифического океанических влияний, а также взаимодействий бореального и центрально-азиатского супераридного воздушных фронтов, представляет собой и своими флорой и растительностью значительный интерес для раскрытия сложных проблем фитогеографии и истории формирования растительного покрова не только юга Сибири и Центральной Азии. К сожалению, флора сосудистых растений пустыни Гоби и Даурии изучена недостаточно, а уж о бриофлоре и лихенобиоте и говорить не приходится. А ведь лихенобиота, особенно эпилитная, по сравнению с флорой сосудистых растений, на порядок более инертна, у лишайников процессы видообразования идут значительно медленнее. Если, изучая структуру флоры сосудистых, мы можем восстановить ее историю за последние 5–10 миллионов лет, то, изучая структуру эпилитной лихенобиоты, мы сможем заглянуть в прошлое на 50–100 миллионов лет. В Гоби эпилитная лихенобиота, по всей вероятности, преемственно развивалась с начала мезозойской эры.
Результаты наших исследований и исследований наших предшественников позволяют говорить о большой оригинальности растительного мира Даурии. Здесь сформировались два своеобразных типа криофильно-аридной растительности – даурские степи-прерии и харганаты (даурская чапараль). В плейстоцене сформировалась формация восточно-сибирских лиственничных лесов из лиственницы даурской (Larix dahurica s.l.) с несколькими подвидами или мелкими видами. В межледниковые эпохи плейстоцена многие неморально-бореальные виды получали возможность возвратиться в те места, откуда они были изгнаны похолоданиями и иссушениями климата. Наличие же на северо-востоке Азии горных цепей меридионального простирания и параллельных им межгорных впадин способствовало проникновению в эпохи похолоданий северных видов на юг, а в эпохи потеплений южных видов на север. Особенно быстро шло расселение южных видов на север в послеледниковые засушливые климатические фазы. Е.В. Вульф (1944) подсчитал количество видов, приведенных В.Л. Комаровым во «Флоре Манчжурии», систематизировал их в ботанико-географические группы и получил при этом следующее распределение манчжурских видов: субтропических и тропических – 128, общих с Японией – 865, общих с Китаем и Монголией – 953, общих с Забайкальем (Даурией) – 788, с Арктической областью – 565, с Охотией – 562, с Европой – 540, с северной Америкой – 341, космополитов – 38, видов эндемичных для Восточной Азии, включая Даурию и Охотский район – 121. Е.В. Вульф пишет, что проникновение тропических и субтропических видов в Манчжурию окончательно угасает и начинается полное господство умеренной флоры, виды которой проникают далеко на запад и на север. Следует заметить, что Е.В. Вульф анализировал флору Манчжурии в широком смысле.
Маньчжурские леса – это своеобразный тип растительности, формировавшийся в течение длительного времени и находящийся в полном соответствии с физико-географическими условиями, довольно устойчивыми, в отличие от резко изменявшегося климата Гоби и Даурии. Но маньчжурский флористический комплекс совмещает неморальные и бореальные элементы, что, несомненно, связано с циклическими изменениями здесь климата то в сторону потепления, то в сторону похолодания. Существенная доля неморально-аридных видов во флоре Манчжурии и Приморья говорит о том, что здесь ритмически изменялась и степень аридности-гумидности климата. Растительность к западу от Большого Хингана в аридные фазы климатического цикла теснее взаимодействовала с флорой Гоби, а в гумидные – с флорой Манчжурии.
Степень аридности в Даурии и Манчжурии не была постоянной, но эти колебания происходили на общем фоне усиливающейся аридизации начиная с конца мелового периода. В результате чего сильно сократилась площадь озер, а пустыни продвинулись на север. Поднятие Большого Хингана также внесло свою коррективу в климат Даурии, сделав его отличным от климата Манчжурии. А.А. Юннатов (1950) считал, что стабильность континентального режима в Центральной Азии способствовала формированию самобытных родов и видов растений, которые могли из Монголии расселяться в соседние страны. Такими растениями он считал некоторые виды караганы, мытника, астрагала, полыни, житняк гребенчатый, вострец китайский, терескен.
В заключение этой статьи можно сделать следующие выводы:
1. Растительный покров – явление весьма динамичное. Рассматривая его в статике, невозможно понять причин возникновения и механизмов формирования его биологического разнообразия. Поэтому изучать растительный покров следует в рамках динамической, а не статической парадигмы.
2. На Дальнем Востоке и в Восточной Азии основными векторами динамики и развития растительного покрова была выработка приспособлений к аридному, гумидному, криоаридному и криогумидному климатам.
3. При общей тенденции усиления аридности климат Дальнего Востока, Центральной и Восточной Азии изменялся ритмически, начиная с палеогена. При этом гумидные фазы сменялись аридными в сложной гармонике колебаний, накладывавшихся друг на друга.
4. Начиная с плиоцена, к общей тенденции усиления аридности климата добавилась тенденция к похолоданию, которая также носила выраженный ритмический характер. При этом потепления и похолодания сменяли друг друга также в сложной гармонике колебаний, накладывавшихся друг на друга.
5. Фазы гумидные и аридные не всегда коррелировали с фазами теплыми и холодными, и это сильно разнообразило климатическую палитру региона.
6. В структуре флоры и растительности Восточной Азии отчетливо видны следы повторяющихся на протяжении миллионов лет аридных и гумидных фаз. В аридные климатические эпохи манчжурские прерии несомненно привлекали кочевников из Даурии, степи и прерии которой в эти эпохи сильно усыхали и превращались в пустыню и полупустыню. В гумидные эпохи в Манчжурии активизировалось земледелие, а в аридные – животноводство.
Литература
1. Беликович А.В., Галанин А.В.. Забайкальский харганат как тип расти-тельности // Комаровские чтения. – Владивосток, 2005. Вып. 52. – С. 98–126.
2. Вульф Е. В. Историческая география растений. – М.: Л.: Изд. АН СССР, 1944. – 545 с.
3. Галанин А.В., Беликович А.В. Даурия как подобласть Манчжурской ботанико-географической области // Комаровские чтения. Владивосток, 2006. Вып. 53. – С. 9–31.
4. Галанин А.В., Беликович А.В., Галанина И.А. Отражение векового климатического ритма в растительности охраняемых территорий российского Дальнего Востока // Доклады международного APN – START Симпозиума по изучению глобальных изменений в Северо-Восточной Азии, Владивосток, 7–8 октября 2002. – Владивосток, 2005. – С. 124–138.
5. Галанин А. В. Гипотеза пульсирующей Земли. – Владивосток: Дальнаука, 2001. –71 с.
6. Галанин А. В. Происхождение таксономического сверхразнообразия растительного покрова (географический и экологический аспекты) // Биоразнообразие экосистем внутреннеей Азии. Т. 1. – Улан-Удэ, 2006. – С. 104–106.
7. Галанин А. В., Беликович А. В., Роенко Е. Н. Конспект дендрофлоры Даурии // Бюллетень Ботанического сада-института ДВО РАН, 2009. Вып. 3. – С. 4–32. Электронный ресурс по адресу: http://botsad.ru /journal/number3/number3_1.pdf
8. Галанин А. В. Ценотическая организация растительного покрова. – Владивосток: ДВО АНСССР, 1989. – 161 с.
9. Галанин А. В. Флора и ландшафтно-экологическая структура растительного покрова. – Владивосток: ДВО АН СССР, 1991. – 272 с.
10. Гумбольдт А. Космос. Опыт физического мироописания. – М.: Мир, 1962. – Ч. 1. – 408 с.
11. Григорьев А. А. О некоторых основных проблемах физической географии // Известия АН СССР. Сер. Геогр., 1957, № 6.
12. Григорьев А. А., Будыко М. И. О периодическом законе географической зональности // Докл. АН СССР. 1956. Т. 110, № 1. – С. 129–132.
13. Намзалов Б. Б. Эндемизм и реликтовые явления во флоре и растительности степных экосистем Байкальской Сибири // Биоразнообразие Байкальской Сибири. – Новосибирск, 1999. – С. 184–192.
14. Наумов И. В. Флора Южно-Чуйского хребта: диссертация ... кандидата биологических наук. – Барнаул, 2009. – 240 с.
15. Попов М. Г. Род Cicer и его виды // Труды по прикладной ботанике, генетике и селекции. 1929 . Т. 21, вып. 1. – С. 1–240.
16. Попов М. Г. Основы флорогенетики. – М: Издательство АН СССР, 1963.
17. Соболевская К.А. основные моменты истории формирования флоры и растительности Тувы с третичного времени // Материалы по истории флоры и растительности СССР. – М.: Л.: изд. АН СССР, 1958. Вып. 3. – С. 249–315.
18. Сочава В. Б. Проблемы физической географии и геоботаники. Избранные труды. – Новосибирск: Наука, Сибирское отделение, 1986. – 343 с.
19. Тахтаджян А.Л. Флористические области Земли. Л.: Наука, 1978. – 247 с.
20. Юннатов А.А. Основные черты растительного покрова Монгольской Народной Республики. / Труды Монгольской комиссии, вып. 39. – М.: Л., изд-во АН СССР – 1950. – С. 1–283.