С целью отграничения Чукотки от Анадырского края по признакам растительного покрова рассмотрим первоначально их совместно, так как история обеих территорий имела одинаковую направленность. Проблема классификации растительности тундры и лесотундры в теоретическом аспекте затрагивалась во многих работах. В этой связи следует особенно упомянуть о работах Ф. В. Самбука (1937), Б. Н. Городкова (1938, 1946), Б. Н. Норина (1958, 1961, 1966, 1979), В. Д. Александровой (1958, 1971, 1964, 1977), Б. А. Тихомирова (1967), В. В. Крючкова (1967) и др. В ряде работ проблема классификации растительности решалась сугубо практически, т. е. описывался растительный покров и основные, составляющие его единицы растительности. Для чукотско- анадырской территории нужно назвать работы Б. Н. Город1933), Л. Н. Тюлиной (1936), В. Н. Васильева (1936, 1956), В. В. Петровского (1967), А. Г. Реутт (1970), А. Е. Катенина (1974, 1981, 1984). Описание растительного покрова с помощью единиц растительности заканчивается обычно его картированием и районированием. Геоботанические районы характеризуются преобладанием какого-либо типа растительности или (и) специфическим сочетанием нескольких типов. Таким образом, тип растительности как классификационная единица является исходным моментом геоботанического и ботанике-географического районирования. К сожалению, типы растительности как таковые на анадырско-чукотской территории не исследовались никем специально в том плане, в котором каждый тип был бы охарактеризован одновременно флористически, биоморфологически, структурально, генетически и экологически. Между тем именно анадырско-чукотскнй регион, возможно, как никакой другой во всей Арктике и Субарктике, имеет особые преимущества для усиленного внимания на классификационную категорию «тип растительности». В этом регионе сочетаются элементы трех природных зон: таежной, лесотундровой и тундровой, имеются разные варианты реликтовой растительности и различные типы вертикальной поясности.

Чукотка. Горный хребет. Акварель Ю.П. Кожевникова
В печати неоднократно обращалось внимание на то, что обычные понятия геоботаники не всегда применимы к тундровой растительности. Полевой опыт свидетельствует о том, что в качестве единиц тундровой растительности следует описывать пятна, выделяющиеся издали одним цветом, хотя бы они и обладали значительными различиями внутри себя. В этом отношении можно заметить, что А. Тэнсли (Tansley) еще в 1920 г. писал, что естественные единицы растительности, от которых мы начинаем, должны, в первую очередь, быть определены эмпирически, без связи с какой-либо теорией. Последовательная ранговая классификация этих «пятен» непременно сталкивается с различными проблемами. Однако отнесение их к тому или иному типу растительности вызывает наименьшие затруднения.
Самое понятие «тип растительности» до сих пор разноречивр, что, впрочем, слабо сказывается на процедуре ботанико-географического деления сравнительно небольших регионов, таких, как чукотско-анадырский. Наиболее уязвимым в этом понятии оказалось применение термина «экобиоморфа», поскольку он объединяет два независимых явления: экоморфу и биоморфу. Было показано, что любая экоморфа включает разные биоморфы, а любая биоморфа — разные экоморфы (Кожевников, 1974, 1978). И. X . Блюменталь (1979) создал новую модель типов растительности с учетом независимого характера экоморф и биоморф. Но практическое использование этой модели, в частности для группы типов микротермных или арктовысотных фитоценозов, по всей вероятности, ограничено, так как многие фитоценозы, принадлежащие одной ассоциации, по режиму увлажнения должны будут относиться к разным типам растительности, что алогично.
Как отметил Б. А. Тихомиров (1947), в Арктике не выработался особый тип растительности, специфический для нее и ей одной свойственный. Высшие таксономические единицы растительного покрова Арктики (типы растительности, формации) повторяют аналогичные единицы других зон. Большинство эдификаторов, широко распространенных в арктической Евразии формаций, являются или видами, связанными теснейшими генетическими узами с горнотундровыми (гольцовыми) областями умеренных и южных широт, или бореальными выходцами из лесов таежной области. Эта точка зрения оспаривается исследователями, признающими существование полярнопустынного типа растительности, выделенного Б. Н. Городковым, который писал в 1947 г., что наиболее древний арктический ландшафт и соответственно тип растительности — полярные пустыни. По его мнению (цит. соч.), климакс полярных пустынь на низменностях представляют моховые полигональные пустыни. Вместо тундровых осоковых в верхнем ярусе преобладают злаки (Alope curus alpinus) и арктическое разнотравье. В. Д. Александрова (1977) отметила, что Городков называл полярными пустынями тундры, а полярнопустынный тип растительности представляют группировки лишайников, печеночников и арктических гипергекистотермных и аркто-альпийских трав. В сущности, Б. Н. Городков понимал полярные пустыни, как тип растительности, аналогично. По нашему мнению, полярные пустыни представляют собой ландшафт, но не тип растительности.
Типы растительности
Мы выделяем типы растительности по доминированию биоморф и по их соотношению в разных ярусах, а также по основному экологическому процессу, если он автокоррелятивен, т. е. направлен на самоподдержание благодаря жизнедеятельности растительности. В качестве примера такого процесса можно назвать самоподдержание болот, которые не обладают специфическими биоморфами, но которые издавна выделяют в качестве типа растительности, а нередко и нескольких. В Анадырско-Чукотском регионе болотный тип выделяется и по соотношению биоморф, среди которых доминируют травянистые гелофиты.
При выделении типов мы руководствовались также представлением о равновесной растительности, о которой Л. Г. Раменский в 1925 г. писал, что равновесной растительности свойственно диффузное сложение, при котором все виды равномерно друг с другом перемешаны. В то же время диффузность является результатом и показателем «напряженной конкуренции растений». Диффузное строение растительности, как известно, очень характерно для тропических лесов, которые из-за их древности естественно оценивать как равновесную растительность и видеть в ее диффузном сложении результат конкуренции. Правомерно распространять эти отношения и на северотаежную, и тундровую растительность, когда в ней существует ценобиоз. Однако диффузное сложение часто характеризует горную, очень разреженную растительность, в которой нет ценотических связей. В таких случаях причины диффузного распределения растений, очевидно, иные. Можно предположить, что в разреженных щебнистых тундрах диффузное распределение растений обусловлено ограниченными тепловыми ресурсами самой поверхности грунта.
Единица поверхности является экраном тепла лишь для малого количества растений, и поэтому они рассеяны равномерно. Нужно иметь в виду, что конкурентная способность растений в любом сообществе связана с фазой его зрелости. Различают 4 фазы: пионерную, строительную, зрелую, дегенеративную (Watt , цит. по: Goldsmith , 1976). В первой и четвертой фазе отдельные растения наименее конкурентоспособны, а в другие «агрессивны». Конкурентоспособность связана и со средним возрастом доминант. Так Calluna на верещатниках живет 35 лет, а в листопадном лесу 200—300 лет (Goldsmith, I 976). В Арктике некоторые зрелые для современных условий сообщества выглядят скорее как пионерные, и только возраст самих доминант свидетельствует об их солидном возрасте. Доминанты эти не являются эдификаторами, т. е. не создают фитосреду. Поэтому несколько спорно утверждение Л. Г. Раменского о том, что обилия растений в установившемся равновесном ценозе строго закономерны. Может быть, для заливных лугов Воронежской губернии это и так, но это далеко не всеобщий вывод. В настоящее время во многих работах показано, что обилие травянистых многолетников часто колеблется год от года, и обусловлено оно погодными условиями текущего года. Так, Т. В. Мальцева (1978) привела данные по приобской лесостепи сухого 1974 г. и влажного 1975 г. Она установила, что в 1975 г. на лугах уменьшилось обилие ксерофитов и увеличилось обилие мезофитов. При этом на некоторых лугах уменьшение было больше (до 10,9%), на других — меньше (4,5%). Автор сделала вывод, что «в более засушливые годы остепненные луга по составу ближе к луговым степям, характеризуются большим участием ксерофитного и мезоксерофильного разнотравья, а в более увлажненные годы они стоят ближе к настоящим лугам за счет увеличения злаковости и мезофитного разнотравья» (1978. С. 15). Характерно, что и сам Л. Г. Раменский отмечал, что в противоположность непрерывно и глубоко сменяющимся количественным отношениям, видовой состав отдельных ценозов относительно устойчив.
Знание закономерностей фаз растительности облегчает идентификацию зрелых сообществ, а также позволяет судить о тенденциях различной растительности после основательного знакомства с нею — стабильна она или сокращает, или расширяет свои позиции, а также о региональных отличиях сообществ одного типа. Так, К. Гимингэм (Gimingham, 1969), изучавший кустарничковые сообщества на Британских островах, нашел, что градиенты среды на континенте повторяются в более океаническом варианте и на этих островах. В результате сообщества с одним и тем же доминантом на континенте и на островах различаются флористически. Полидоминантные кустарничковые сообщества различаются по соотношению покрытий доминантов. Классификация по соотношению покрытий доминантов увязывается с флористической классификацией. Однако континуальный характер изменений сообществ как в отношении покрытий, так и флористического состава позволили К. Гимингэму выделить наряду со стабилизированными, зрелыми сообществами целый ряд переходных, не являющихся единицами классификации. Такое решение проблемы нельзя назвать удачным, так как любая классификация должна вмещать все разнообразие классифицируемых объектов. Поэтому возникает вопрос — как быть с сообществами, не относящимися к фазе зрелых? Ведь как справедливо отметил Л. Г. Раменский (1925), если материал обширен и полон, то выявляется бесконечное разнообразие слабо и резко различных растительных группировок, а объем сочетаемости каждого вида растений с другими велик и неповторяемо своеобразен. Действительно, нет таких сообществ, которые бы в точности совпадали по всем своим показателям, как нет абсолютно сходных растений. Но отнесение любого сообщества в любой фазе его развития к тому или иному типу растительности легче, чем к той или иной ассоциации.
Уместно провести параллель с определением неизвестных нам растений. Определение мы начинаем с выяснения отдела (или класса), затем семейства, рода и вида, т. е. идем сверху вниз. Могут возразить, что в таксономии растений эта процедура естественна, так как имеется система, а в геоботанике нет системы. На это можно ответить, что и в таксономии растений существуют свои разногласия, имеется несколько систем растений, но оттого, спереди или сзади системы помещаются однодольные, определение растений не зависит. В геоботанике разработана система таксономических единиц, имеющих определения, которыми мы и пользуемся. Другое дело, что одни исследователи принимают одно число региональных структур растительности, а другие другое, но такая же ситуация существует и в таксономии растений. Это говорит лишь о необходимости дальнейших методологических исследований.
Представление о характере равновесной растительности данной местности складывается не только при наблюдении ее диффузного строения, но и из факта высокой частоты встречаемости участков этой растительности, а также из площади этих участков, откуда возникло понятие минимум-ареала растительности. Это понятие используется как концепция и как характеристика растительности, но дает больше неопределенности, чем ясности (Goldsmith , 1976). При определении минимум-ареала базируются на одном из трех критериев: флористическая структура, частота видов и гомогенность. Мы принимали во внимание только гомогенность, полагая, что при морфологическом подходе к растительности это наиболее существенно, тогда как флористические признаки важны при определении динамики растительности.
Таким образом, в основу выделения типов растительности нами положен эколого-физиономический принцип. Использование этого принципа при выделении типов растительности избавляет исследователей от многих забот, в том числе и определения растений. Этот принцип использован П. Дансеро (Dansereau, 1956, 1957, Dansereau et al., 1966) при создании им универсальной системы описания растительности. Его система основана на 6 признаках: биоморфа (habitform ), форма и размеры листа, текстура листа, сезонность, ярусность, покрытие, П. Дансеро подчеркивал необходимость описания растительности в понятиях, касающихся только ее. Этой же точки зрения придерживался другой сторонник физиономического описания А. Кюхлер, по мнению которого флористическая структура и экологические аспекты растительности исследуются после того, как изучены жизненные формы и структура растительных сообществ, т. е, их физиономия (Kuchler, 1951). Д. Найт (Knight, 1965) разработал структурно-Функциональный индекс для характеристики растительности прерии в Висконсине. Он подчеркнул, что его индекс может быть широко использован вне зависимости от флористической структуры растительности.
Эколого-физиономический подход к выделению типов растительности, на наш взгляд, наиболее оправдан в адекватном отображении современной природной дифференциации региона. Он подготавливает основу для дальнейшего историко-флористического и экологического анализа. При первой попытке выделения типов растительности Анадырско-Чукотского региона было признано существование 13 ее типов (Кожевников, 1976). Ознакомившись позднее с типами растительности, выделенными В. Д. Александровой (1977), я объединил некоторые свои типы, следуя этому автору, одновременно признав, что некоторые ее типы очень условны, например, стланиковый, два болотных, полярнопустынный. В настоящее время в Анадырско-Чукотской области нами различаются следующие типы растительности.
Лесной тип
К этому типу относятся классы формаций хвойных и лиственных лесов, распространенных в основном в бас сейне Анадыря. Хвойные леса включают лиственничники и кедрово-стланиковые леса. Формации кедрового стланика, как это рекомендовано В. Б. Сочавой (1956), Б. П. Колесниковым (1961) и др., следует рассматривать в качестве лесных, а не кустарниковых, как считает Т. Г. Дервиз-Соколова (1967в) и другие авторы. В плакорных условиях Марковской впадины кедровый стланик нередко имеет форму роста «многоствольное дерево» с восходящими стволами (Кожевников, 1978). Заросли кедрового стланика являются каналом проникновения на север бореальных компонентов биоты. Признание формаций кедрового стланика как лесного типа растительности (Пивник, 1958) позволяет очень логично оконтурить зону лесотундровых ландшафтов, на что еще в 1939 г. обращал внимание зоогеограф Л. А. Портенко и что нашло отражение на «Карте геоботанического районирования СССР» (1947). Северная граница берингийской (кустарниковой) лесотундровой области показана на этой карте чрезвычайно точно.
|

Лиственичное редколесье. Фото А.А. Галанина |
Лиственничники распространены в верховьях Анадыря, где они почти смыкаются с чаунскими лиственничниками (Тюлина, 1936) (согласно карте типов растительности, составленной А. Г. Реутт (1970), они там смыкаются и в среднем течении Майна. По указанию А. П. Васьковского (1958), лиственничники в верховьях Анадыря занимают почти островное положение, но все же соединяются с анюйскими лесами по р. Яблон. «Изолированный» массив лиственницы в долине Майна имеет площадь 1 млн. га (Реутт, 1970).
В. Н. Васильев (1956) понимает под лесотундрой сочетание лесных и безлесных участков одного ландшафта и называет лесотундрой весь район среднего течения Майна, однако в одном месте своей работы (с. 157) он явно имеет в виду, что долинные лиственничники представляют все же лесной тип растительности. Так он пишет: «Данные лиственничники (вдоль рек, Ю. К.) быстро переходят через лесотундру (разрядка моя, Ю. К.) с низкорослой корявой лиственницей, с крайне небольшой полнотой насаждений, в тундру». Ниже еще будет сказано о «тундре» в бассейне среднего течения Майна. Флористически лиственничники и кедровники в бассейне среднего Анадыря неспецифичны, заслуживают внимания лишь присутствием в них Сагех globularis, Ledum palustre, и обилием Betula middendorffii. В наиболее продуктивных лиственничниках на склонах небольших возвышений деревья достигают высоты 15—18 м и диаметра 15—25 (35) см. В них обычен кустарниковый подлесок из Salix krylovii, S . pulchra var. anadyrensis, Betula nana ssp. exilis, B. middendorffii. В напочвенном покрове обычны Vaccinium uliginosum, Rubus arcticus, Ledum palustre, Pyrola rotundifolia ssp. incarnata , Calamagrostis purpurea ssp. langsdorffii и др. |

Среднее течение реки Белой. В пойме чозениевые леса перемежаяюся с высокими ивняками и ольховниками |
Лиственничники на надпойменных террасах (высотой около 8-10 м) с диаметром деревьев до 20 см. В их кустарниковом подлеске обычны Ribes triste, Alnus fruticosa ssp. kamtschatica и Rosa acicularis. Еще Л. Н. Тюлина (1936) отмечала, что на Майне и Алгане лиственница повсеместно распространена на сопках, образуя там полосу корявого суховершинного редколесья шириной не менее 10—12 км в обе стороны от рек. По нашим наблюдениям, в виде стланика лиственница встречается вместе с ольхой и на плоских вершинах порядка 300 м. Pinus pumila, распростертый по щебню, здесь довольно редок. Betula platyphylla ssp. minutifolia и Sorbus anadyrensis поднимаются по южным склонам до 150—200 м.
В бассейне среднего Анадыря широко представлены чозенники, тополевники, березняки, ольховые (Alnus hirsuta) и ивовые (Salix udensis, S. schwerinii) леса, все, кроме березняков, пойменные. Березняки из Betula platyphylla ssp. minutifolia, вопреки установившемуся взгляду (Стариков, 1958), не являются пойменными лесами, встречаясь исключительно на незаливаемых террасах и высоких увалах; в бассейне Майна есть горные березняки, внешне очень сходные с каменными березняками из Betula lanata в северной Охотии. |
Пойменные леса часто полусумрачны в глубине, где напочвенный покров нередко состоит из одного хвоща (Equisetum arvense). По опушкам в разреженных лесах развивается крупный травостой из Lactuca sibirica, Anthriscus aemula, Aruncus kamtschaticus, Cacalia hastata, Thalictrum minus ssp. kemense, Urtica angustifolia и др., с кустами Ribes triste и R. dikuscha, на Майне — Lonicera edulis, Rubus sachalinensis.
Чередуемость лиственничников и лиственных лесов обусловлена, по-видимому, мощностью слоя супеси и суглинка на надпойменных террасах. Там, где этот слой 0,5 м и более, расположены лиственничники, а если он тонкий, то на нем развиты лиственные леса. Под слоем супеси и суглинка, обычно оторфованом или пропитанном гумусом, залегает галечная толща, в которой циркулируют грунтовые воды, препятствуя подъему зеркала вечной мерзлоты.
Луговой тип
Настоящие луга не выходят за пределы распространения лесного типа растительности. Большинство лугов антропогенны, но встречаются луга и естественного происхождения, особенно на склонах в долинах и на надпойменных террасах. Склоновые луга отличаются высокотравьем, несколько напоминая луга Камчатки. Обычными видами на них являются Geranium erianthum, Aconitum delphinifolium s. 1., Chamaerion angustifolium, Veratrum oxysepalum, Tanacetum vulgare ssp. boreale, Ptarmica alpina и др. Близ старых поселков обычны закустаренные луга, преимущественно злаковые. На них растут Elymus confusus, Е. borealis, Arctagrostis arundinacea, Agrostis anadyrensis, Bromopsis pumpelliana, Poa alpigena, Trisetum spicatum, Hordeum jubatum, Artemisia tilesii, Polygonum aviculare s. I., Lactuca sibirica, Aster sibiricus, Rumex sibiricus и др. Видовой состав лугов довольно беден для этого типа растительности. Распределение видов обычно очень равномерное.
Начальные стадии залуговения хорошо заметны на внешних кромках огромных приречных отмелей. Склоны террас проток покрыты также луговой растительностью. Можно сказать, что близ рек все участки, не имеющие по каким-либо причинам древесно-кустарниковой растительности, заняты луговой растительностью, но в удалении от речных систем, включая многочисленные протоки, луговая растительность практически отсутствует.
Лесотундровый тип
Особенности этого типа подробно освещены Б. Н. Нориным (1961). Это редколесья на эрикоиднокустарничково-моховом или ерниково-моховом напочвенном покрове. К этому типу относятся также сильно разреженные заросли кедрового стланника. Как указал Б. А. Тихомиров (1967), «главнейшее отличие лесотундровых сообществ от северотаежных лесов — расстройство ценотического единства, или утрата эдификаторной роли древесных пород». В лесах альбедо ниже, чем в тундрах. Это позволяет лесу получать больше тепла (Tuhkanen, 1980). Следовательно, лес обладает свойством автокорреляции. В лесотундровых редколесьях это свойство в значительной мере уменьшается, но, очевидно, не утрачивается совсем. Редколесья практически не оказывают влияния на растительность нижних ярусов, представляя яркое свидетельство явления инкумбации — наложения ярусов, описанного В. Б. Сочавой (1932, 1933) как раз в Анадырском крае.
Основная единица растительности в лесотундре — комплекс синузий, часто сильно различных по экологии даже в одном ярусе (Нории, 1967, 1979). Для нижних ярусов лесотундровых лиственничных редколесий характерны кедровый стланик, ольховник, березка Миддендорфа, Сагех soczaveana, Eriophorum vaginatum, Salix myrtilloides, багульник, голубика, подбел, клюква мелкоплодная и др. Моховую дернину слагают преимущественно сфагновые мхи.
Кустарниковый тип
Этот тип слагают ерники из Betula middendorffii и В . nаnа ssp. exilis с ортотропным ростом в континентальных районах, ольшатники из Alnus fruticosa s. str. и ssp. kamtschatica (последняя раса в континентальных районах ), ивняки из Salix krylovii, S. alaxensis, S. lanata ssp. richardsonii, S. glauca, S. pulchra s. str. и var. anadyrensis, в континентальных районах — S. schwerinii. Напочвенный покров может быть самым различным — от галечникового разнотравья до болотной дернины с обилием пушицы, Сагех lugens и эрикоидных кустарничков.
На Чукотке основными кустарниковыми формациями являют ся ольшаники на склонах гор и ивняки в поймах рек. Ольшаники распространены в основном на материковой Чукотке, но отдельные массивы их имеются и на востоке Чукотского полусотрова, распространяясь на север вплоть до оз. Коолень. Видовой состав ольшаников весьма постоянен из-за наличия более или менее развитого мохового покрова. Он включает обычно Vaccinium uliginosum, Betula папа ssp. exilis, Ledum palustre ssp. decumbens, Polygonum tripterocarpum, Spiraea stevenii, Artemisia tilesii, Calamagrostis purpurea ssp. langsdorffii, Rubus chamaemorus, Lycopodium annotinum ssp. pungens; на юге Чукотки в ольшаниках обычен Rhododendron aureum. В нижнем ярусе доминировать может любой из первых трех вышеназванных кустарничков.
Пойменные ивняки распространены почти по всей Чукотке, за исключением северных приморских равнинных районов. Крупные ивняки на Чукотском полусотрове на р. Уттавеем только 40 км не достигают северного побережья. Основным фактором их существования является укрытие зимой снегом и относительно быстрое стаивание снега весной. Поэтому в океанических и приморских районах они приурочены к достаточно врезанным долинам речек и их высота строго соответствует глубине долины. Ветви, торчащие из-под снега, быстро отмирают. Пойменные ивняки, как правило, разнотравные. В их наборе видов обычно принимают участие Aster sibiricus, Cardamine hyperborea, Valeriana capitata, Polemonium acutiflorum, Calamagrostis purpurea ssp. langsdorffii, Сагех podocarpa, Astragalus alpinus и др. Склоновые ивняки, весьма обычные в континентальных районах Чукотки, в океанических ее районах распространены меньше, и площади, занимаемые отдельными массивами, невелики. Они приурочены к различным укрытым местоположениям или, наоборот, к выступам в нижних частях склонов, под которыми нередко лежат снежники. Склоновые ивняки также преимущественно травяные, но их видовой состав очень непостоянен. Доминантами в этих ивняках могут быть Salix krylovii, S . lanata ssp. richardsonii, S . pulchra , S. glauca.
В континентальных районах березка тощая, голубика и багульник нередко образуют низкокустарниковые заросли до 0,5 м высотой. Эти заросли заполняют большинство благоприятных местообитаний, тем самым обедняя флору.
Болотный тип
В рамках этого типа выделяется два подтипа: пресный болотный и лайдовый (приморские засоленные болота). В лесных районах бассейна Анадыря роль болот в ландшафтах чрезвычайно велика, несмотря на резко континентальный климат. Они тянутся беспрерывно на многие километры, хотя растительность на этом пространстве нередко существенно изменяется: участки чистых кочкарников покрываются зарослями кустарников, которые переходят в лиственничные редколесья или сменяются зарослями кедрового стланика, не уступающими зарослям на склонах гор. Обычный состав видов включает Carex aquatilis ssp. stans, С. limosa, Eriophorum vaginatum, Betula nana ssp. exilis, Chamaedaphne calyculata, Ledum palustre, Vaccinium uliginosum, Andromeda polifolia, Rubus chamaemorus, Pedicularis lapponica. Обычны кусты и мелкие деревца лиственницы.
Болотообразование интенсивно происходит и в наши дни в связи с тем, что в бассейне Анадыря существуют оптимальные условия для роста сфагновых мхов, удерживающих влагу. Заболачивание начинается также там, где происходит нарушение целостности зеленомошной дернины и вечная мерзлота подтаивает. Это происходит, в частности, на звериных тропах. Особый тип болот представляют затянутые сплавиной старицы. Их флористический состав очень постоянен. Он включает Carex rhynchophysa, С. lapponica, С. chordorrhiza, С. limosa, Comarum palustre, Eriophorum polystachyon, Cicuta virosa, Equisetum fluviatile, Arctagrostis arundinacea, Chamaedaphne calyculata, Salix myrtilloides, S. pulchra, AInus fruticosa.
Образование сплавины по краям низинных и старичных озер очень обычный процесс в бассейне Анадыря. Он может быть прослежен в разных стадиях развития. В тундровой зоне сплавинообразование можно встретить лишь на южных приморских низинах, например, на восточном берегу зал. Креста, где на сплавинах озер массовы Hippuris vulgaris, Arctophila fulva, Ranunculus pallasii, Carex aquatilis. Весьма характерно, что болота с высокорослыми осоками С. rhynchophysa, С. vesicata, С. rostrata и набором видов, перечисленных выше, распространяются на север дальше границы леса в бассейне Анадыря. Они встречаются, например, в низовьях р. Белой (притока Анадыря). Однако собственно на Чукотке этих болот уже нет. Кроме того в анадырском крае имеется серия монодоминантных болот, отсутствующих на Чукотке: хвощевые (Equisetum fluviatile), веховые (Cicuta virosa), осоковые (с Carex rhynchophysa и С. vesicata) и др. Для этих же районов характерны сильно высыхающие болота, называемые обычно тундрой.

Сочетание осоково-гипнового болота и кустарничковой тундры.
Фото А.В. Беликович |
Чукотские болота очень разнообразны, поскольку их формированию способствует близость зеркала вечной мерзлоты и превышение количества осадков над испарением. Процесс болотообразования автокоррелятивен, т. е., возникнув, очаг заболачивания, как правило, не деградирует, а развивается, причем, чем далее, тем интенсивнее. Имеются болота торфяные и без торфа, но с постоянным избыточным увлажнением, олиготрофные и эутрофные, сфагновые и гипновые, с морозообойными пятнами и без таковых, бугристые, бугристомочажинные и ровные. Очевидно, с болотами не следует смешивать кустарничково-моховые тундры с торфянистыми почвами мощностью до 20 см, которые, будучи мокрыми, напоминают болота. Различительными признаками тундр и болот является наличие в последних торфяного и глеевого горизонтов в одних случаях и постоянная избыточная увлажненность в других, например, в пойменных болотах. Однако в ряде случаев возникают затруднения — относить ли данный участок к тундре или рассматривать его как болото. Очевидно, это связано с начальными стадиями заболачивания, которое представляет на современном этапе весьма интенсивный процесс естественного развития ландшафтов Северо-Востока. Последнее обусловлено избытком осадков над испарением; коэффициент увлажнения, по Н. К. Клюкину (1970), на всем азиатском Северо- Востоке составляет 1,33, т. е. 25% выпавших осадков не испаряются. |
Болота обнаруживаются на различной высоте, отсутствуя лишь на вершинах гор. Даже на привершинных террасах гор до 600 м высоты они вполне обычны. На привершинной слегка наклонной террасе горы Амгуэмы существует обширное бугристое кустарничково-осоково-моховое болото с редкими мочажинами. Его видовой состав весьма типичен для болот на подобных местоположениях. Он включает Сагех bigelowii ssp. lugens, Betula nana ssp. exilis, Eriophrum vaginatum, E. polystachyon, Salix pulchra, S. fuscescens, Vaccinium uliginosum ssp. microphyllum, Ledum palustre ssp. decumbens, Luzula wahlenbergii, Pedicularis lapponica, P. albolabiata, Hierochloe alpina, Diapensia lapponica ssp. obovata.
К болотному типу относятся и кочкарники из Eriophortim vaginatum, хотя многие из них довольно сухие. Кочкарники встречаются сфагновые и зеленомошные. Многие из них характеризуются значительной ролью Betula nana ssp. exilis, которая встречается почти на всех болотах, за исключением северных приморских. Сагех aquatilis ssp. stans является одним из основных болотных ценозообразователей. Ее заросли желтоватого цвета уже издалека отмечают осоковые болота. Эти болота располагаются на пологих склонах, на днищах равнин, на надпойменных террасах, вокруг озер. Иногда в них велика роль Salix pulchra, кустики которой не превышают стеблей осоки. Видовой состав болот на ровных участках весьма однообразен. Наиболее постоянные виды, кроме названных выше, это Andromeda polifolia, Carex vaginata, С. rotundata, Calamagrostis lapponica, Polygonum tripterocarpum, Pedicularis parviflora ssp. pennellii.
На эутрофных болотах, чаще расположенных на шлейфах, видовой состав значительно обогащен. Здесь встречаются Salix reticulata, Claytonia tuberosa, Eutrema edwardsii, Pedicularis capitata, Carex scirpoidea, C. misandra, C. ledebouriana, C. fusci - dula, Senecio kjellmannii, Tofieldia coccinea, Anemone sibirica, Beckwithia chamissonis и др. Эутрофность болот обусловлена миграцией элементов минерального питания с выше расположенных участков склонов гор, когда они образованы основными или щелочными породами. Водное питание болот на шлейфах осуществляется в значительной мере при подтаивании вечной мерзлоты на склонах гор. Это позволяет болотной дернине нарастать вверх по склону. Наряду с этим возможны случаи, когда обогащенный видовой состав болота обусловлен нейтральностью среды (при отсутствии бокового подтока грунтовых вод, например, на поверхностях надпойменных террас, со всех сторон окруженных протоками). Как отмечалось ранее (Кожевников, 1976, 1978, 1979), на таких участках не задерживаются ни основные соли, ни кислые продукты, в результате чего реакция почв нейтральна.
На шлейфах и на надпойменных террасах нередки бугристые болота с галечниковым субстратом между буграми, более или менее сырым, иногда залитым водой проточной или полузастойной. Сфагновые болота более распространены в континентальных районах, чем в океанических, где сфагны чаще имеют вид небольших латок. Еще Б. Н. Городков (1916) отметил, что сфагновые болота более свойственны лесной зоне, а гипновые (в широком смысле) тундре и степи. Теперь нам известно, что в подзоне, кустарниковых тундр сфагновые покрытия занимают большие площади. К сфагнам тесно привязаны такие континентальные виды, как Pinguicula spathulata, Oxycoccus microcarpus. Наличие сфагновых участков в океанических районах можно объяснить их реликтовым состоянием.
В континентальных районах болота нередко закустарены ивами и ольхой. Alnus fruticosa может образовывать на них ярус от разреженных низких кустиков до почти сомкнутых насаждений до 2—2,5 м высотой. На бугристо-мочажинных, бугристых, валиковых болотах обильны гипарктические кустарнички, Salix pulchra и S. fuscescens. Несмотря на интенсивное болотообразование на Чукотке, в континентальных ее районах и реже в океанических некоторые болота, особенно с тонким торфяным слоем подсыхают. Обычно это связано с неотектоническими поднятиями, в результате которых увеличивается сток. На таких болотах обычно доминирует Carex bigelowii ssp. lugens, иногда массовы Loiseleuria procumbens, Claytonia acutifolia, Novosieversia glacialis, Polygonum bistorta ssp. ellipticum и др., обильны лишайники.
|

Гнилой Угол Анадырского залива. Лайды. Фото Д.А. Галанина |
Лайдовый подтип — это растительность засоленных тяжелых субстратов морских побережий, называемых также маршевыми или тамповыми болотами. Исходя из эколого-физиономического принципа выделения типов растительности, лайдовый подтип не может быть включен в луговой, как это принимал А. П. Шенников (1938). Этот автор дал привычное теперь определение луговой растительности, как сообществ травянистых многолетних мезофитов и не признавал за луговую растительность сообщества ксерофитов (типчака, кобрезии), но с другой стороны по отношению к фактору увлажнения он не сделал ограничения луговой растительности и принимал за таковую сообщества гигрофитов. Лайдовый подтип представляет гигро- и гидрофитная галофитная растительность, в которой доминируют Carex subspathaсеа, Calamagrostis deschampsioides, Puccinellia tenella, P. phryganodes. Для этой растительности характерны также Cochlearia arctica, Dendrathema hultemii, Honkenya peploides, Potentilla egedii и др.
|
В. Д. Александрова (1977) отнесла приморские галофильные болота к травяногипновым гомогенным тундровым болотам также в качестве подтипа. Однако она выделила в качестве самостоятельного типа бугристые и полигональные комплексные тундровые болота, которые, на наш взгляд, не могут быть противопоставлены другим болотам иначе как по микрорельефу. Лайдовый болотный подтип на Чукотке довольно часто занимает места с бугристым или валиковым микрорельефом и большим количеством озерков. Отделить бугристые лайды от ровных по геоботаническим признакам невозможно, и это говорит лишний раз о единстве болотного типа растительности. Б. А. Тихомиров (1945), как и А. П. Шенников (1938), рассматривал лайды, как арктические приморские луга с наиболее распространенной ассоциацией Сагех subspathacea, Puccinellia phryganodes с участием Stellaria humifusa. Он считал (цит. соч.), что существуют луга из мезофитных галофитов. По-видимому, такие луга не являются коренной (климаксной) растительностью.
Тундровый тип
Этот тип включает тундры, называемые типичными, т. е. моховые, точнее кустарничково-моховые, с большим или меньшим обилием осоковых. Причем здесь имеются в виду именно кустарничково-моховые сообщества, а, допустим, не пятнистые тундры, которые, как отметил Б. Н. Норин (1966), являются определенной категорией при классификации растительного покрова, но не растительности. То же самое относится к полигональным тундрам. Аналогичным образом можно рассматривать и куртинные парковые заросли кустарников, т. е. между куртинами кустарничково-моховая растительность относится фитоценотически к тундровому типу. Обычно она не отличается от подобной растительности за пределами кустарникового массива.
|

Кустарничковые тундры на высокой надпойменной террасе.
Фото А.В. Галанина |
К числу доминантов этого типа относятся Dryas octopetala s. L, D. integrifolia (на известняках), Cassiope tetragona, Vaccinium uliginosum ssp. microphyllum, V. vitis-idaea ssp. minus, Ledum palustre ssp. decumbens, Betula nana ssp. exilis, Empetrum nigrum, Arctostaphylos alpina, Carex bigelowii ssp. lugens. Каждый из этих видов может доминировать индивидуально или в смеси с другими видами (полидоминантные тундры). На влажных склонах широко распространены кустарничково-осоково- моховые тундры, в которых большая роль принадлежит Сагех bigelowii ssp. lugens, но обычны также С. scirpoidea, С. vaginata, С. nesophila (на восточной Чукотке), С. podocarpa; из кустарничков здесь обычно представлены дриада (фоновое), Rhododendron parvifolium, Cassiope tetragona, Diapensia lapponica ssp. obovata; часто имеются редкие кустики Salix glauca и S. pulchra. Разнотравье очень непостоянно. Особенно характерными видами являются Luzula tundricola, L. nivalis, Hierochloe alpina, Saussurea tilesii, Novosieversia glacialis, Saxifraga nelsoniana, Ciaytonia acutifolia, Pedicularis capitata, Polygonum bistorta ssp. ellipticum, Poa arctica, Hedysarum hedysaroides ssp. arcticum, Anemone sibirica, Eutrema edwardsii, Nardosmia glacialis, Astragalus umbellatus. Как уже было сказано, болотный тип отличается от тундрового не по признакам растительности, которая в ряде случаев может быть идентичной. Обводненность кустарничково-моховых сообществ, как правило, временная, но иногда весьма постоянная за счет грунтового притока вод от подтаивания мерзлоты или снежников на более высоких уровнях. Как и в болотах, это приводит к снижению кислотности и как следствие — к флористическому обогащению. |
В таких тундрах на небольшой площади обнаруживается до 30 и более видов, среди них Сагех fuliginosa ssp. misandra, С. vaginata, С. ledebouriana, Luzula wahlenbergii, Oxytropis maydelliana, Thalictrum alpinum, Androsace bungeana, Senecio atropurpureus, S. frigidus, Salix reticulata, Pedicularis oederi, P. albolabiata, Gentiana algida, Eutrema edwardsii и др. Co временем такие участки заболачиваются. На дренированных участках шлейфов с богатым минеральным питанием некоторые тундры настолько насыщены разнотравьем, что имеют характер переходный к луговинам.
Растительность тундрового типа занимает различные местоположения от низких надпойменных террас до плоских вершин низких гор или седловин между острыми вершинами. Однако тундры с доминированием вороники, голубики и брусники не поднимаются выше шлейфов гор. В этих тундрах часто обнаруживаются бореальные виды Pyrola minor и Linnaea borealis. В эутрофных кустарничково-осоково-моховых тундрах на известняках с Dryas integrifolia часто содоминирует Salix rotundifolia, а видовой состав насыщен кальцефилами и базифилами: Artemisia globularia, Carex membranacea, Ciaytonia tuberosa, Potentilla biflora, Silene acaulis и др.
Степоидный тип
Под этим типом понимается ксероморфная разреженная растительность каменистых и щебнистых участков склонов. Ценотические отношения в этой растительности отсутствуют, и флористический состав подвержен сильным колебаниям. В этот состав входят виды: 1) в более континентальных и южных областях являющиеся по преимуществу степняками (Ptilsatilla nuttalliana ssp. multifida, Helictotrichon krylovii и др.); 2) тяготеющие к степоидам лишь на Чукотке, а в других областях встречающиеся и вне обстановки остепнения (Lychnis sibirica, Silene repens и др.); 3) на протяжении всего ареала не избегающие условий остепнения, но и не привязанные строго к ним (Dracocephalum palmatum, Festuca lenensis и др.). Уже в пределах Чукотского национального округа разные «степняки» ведут себя по-разному. На Чукотке они могут быть верными компонентами степоидов, а в Анадырском крае обитать в составе криоксерофитной или какой-либо иной растительности. Lychnis sibirica, например, встречается в Анадырском крае на редкотравных щебнистых склонах гор, где других «степняков» нет. Степоиды только тем и характеризуются, что часть видов их составляющих, в более южных районах обитает в степях, т. е. выделяются они скорее флористически, чем геоботанически, потому что на Чукотке имеются группировки по набору биоморф очень сходные с «остепненными», но не называемые так, например, участки с доминированием Carex rupestris, Festuca altaica или Poa glauca. Многие примечательные чукотские «степняки» в других районах своего ареала расцениваются ботаниками без всякой связи со степями. В Карпатах Сагех pediformis назван «чернолесным восточным видом» (Попов, 1949). А. И. Толмачев (1938) относил Thymus serpyllum к бореальному элементу, такому как Arctostaphylos uvaursi. И действительно, оба эти вида являются боровыми. В 100 км к югу от пос. Барроу на Аляске Carex obtusata обитает на свежих речных аллювиях, выступая вместе с Elymus arenarius пионерным задернителем песков (Peterson, Billings, 1980).
Понятие «степоиды» было предложено В. Б. Сочавой (Сочава, Липатова, 1960) для травяных группировок ксерофитов и мезо-ксерофитов в амурской подтайге. Оно призвано подчеркнуть некоторое подобие этих группировок степным фитоценозам. В этом же смысле мы неоднократно использовали этот термин для Чукотки, поскольку понятие «степь» к обсуждаемой растительности неприменимо, хотя бы и сопровождаемое эпитетом «каменистая» или «субарктическая». Уже на расстоянии нескольких метров эта растительность теряется из виду и неслучайно Б. Н. Городков (1952) трактовал «арктические степи» как пионерную стадию растительности свежих ледниковых наносов. Согласно В. Б. Сочаве (Сочава, Липатова, 1960), степоиды представляют динамическую серию ассоциаций «с лесом в конце». Это не коренная растительность, а вторичная. Их залесение без вмешательства человека (или пожара) обеспечено.
Таким образом, если в понятие степоид вкладывать узкое региональное значение, то Б. А. Юрцев (1978) прав, когда заявляет, что мое перенесение этого понятия на аналогичную растительность Чукотки неправомерно, не соответствует смыслу, вложенному в это понятие В. Б. Сочавой. Однако понятие это касается растительности, а не видового состава. Поэтому оно может быть распространено на любую аналогичную в плане строения растительность. Ряды степоидов «с лесом в конце» характерны для разных регионов. Они были, например, описаны нами на Индигирке и на Колыме (Кожевников, 1981). Североякутские степоиды по всем признакам не отличаются от чукотских, за исключением того, что у последних не может быть «леса в конце». Следует заметить, что в районах, где имеется лесная растительность, степоиды на Чукотке выражены лучше, и то, что они встречаются и в абсолютно безлесных районах, свидетельствует лишь об их лучшей сохраняемости при изменениях климата, так как это сугубо околоземная растительность. В бассейне Анадыря ряды степоидов имеют «лес в конце», в том числе заросли кедрового стланика, которые вытесняют степоиды (Кожевников, 1977а).
|

На южных склонах низких гор в континентальных районах на участках КДЛЭ (светлые пятна) существуют многоликие степоиды. На переднем плане типичная тундра с отдельными низкими кустами ив в условиях ГЛЭ. Фото Ю.П. Кожевникова |
По данным Б. А. Юрцева (1967), все местонахождения «степных сообществ на северо-востоке СССР находятся к северу от ареалов лиственницы и кедрового стланика и в гольцово-таежных ландшафтах бассейна Анюя они отсутствуют из-за того, что их экотопы — сухие южные склоны заняты зарослями, кедрового стланика. Теперь известно, что степоиды широко представлены в подзоне северной тайги и дизъюнкция в их распределении, показанная Юрцевым (1974, рис. 3), не существует. По-видимому, мысль об этой дизъюнкции и давала повод думать, что «степные сообщества» тундровой зоны представляют собой нечто самобытное, отличное от более южных степей и им подобной растительности. Таким образом, если не принимать термин «степоид» по отношению к анадырско-чукотским участкам разреженных относительных термоксерофитов, то нужно вводить какойто новый термин, в чем нет особой необходимости.
|
Проблема обозначения горной ксерофитной растительности обсуждалась многими исследователями в разных регионах. А. А. Гроссгейм (1948) использовал для обозначения разреженной ксерофитной растительности Кавказа термин «нагорноксерофильная», впрочем, называя его неудачным. Этот большой знаток растительности Кавказа считал, что понятие «нагорно-ксерофильная степь» не может быть использовано для нагорно-ксерофильной растительности. Он пишет: «Термин степь тем более не может быть принят для типа нагорно-ксерофильной расти-тельности уже потому, что дерновые процессы ей по существу чужды и если и развиваются, то не как связный процесс на целостной территории, а как фрагменты дерновых микрогруппировок, в особых микроусловиях (Гроссгейм, 1948. С. 88). А. А. Гроссгейм различает гаммаду, фригану и шибляк, определяемые по жизненным формам.
Характеризуя растительный покров Алтая, Г. Н. Огуреева (1980. С. 19) пишет, что «вероятно, следует отказаться от широко употребляемого, но неудачного из-за своей неопределенности названия «каменистая степь». С автором можно согласиться и понятие «степоид» вполне заменяет понятие «каменистая степь», которое означает по сути своей не сомкнутую растительность, а со значительной ролью обнаженного каменистого субстрата. На склонах надпойменных террас степоиды часто расположены не на каменистом, а на суглинистом или супесчаном субстрате.
В Гренландии аналогичные «степные» участки приурочены к лессовым почвам ( Bocher , 1949, 1954). Однако лёсс, по Т. Бехеру — это ледниковая грязь, выносимая реками в половодье, которая затем оседает, высыхает и разносится ветром. Такие «лёссы» широко распространены в бассейне Анадыря. По существу, эти отложения являются не лессами, а лессовидными суглинками. П. И. Полевой (1915) писал, что среди континентальных отложений наиболее широкое распространение имеют иловатые тонкозернистые отложения, напоминающие лёсс. Эти отложения представляют собой тончайший продукт разрушения пород, перемешанный с растительными остатками. Ил имеет способность, подобно лёссу, образовывать отвесные стенки в обрывах террас и покрывает грубозернистые отложения.
Судя по описаниям Т. Бёхера ( Bocher , 1954, 1963), в Гренландии нет настоящей степной растительности, а имеются лишь степоиды, нередко представляющие пятна среди кустарничковой растительности с Betula папа и Ledum palustre ssp. decumbens. Бёхер и сам называет эти «пятачки» степоидов steppelike patches. Carex supina ssp . spaniocarpa — один из доминантных видов степоидов Т. Бёхер называет низкоарктическим континентальным ксерофитом. Типичным степным сообществом в Зап. Гренландии Т. Бёхер ( Bocher, 1963) считает следующее с доминированием Carex supina ssp. spaniocarpa на южном склоне с крутизной 10°: Calarnagrostis purpurascens, Carex rupestris, Artemisia borealis. Campanula giseckiana, Cerastium alpinum ssp. lanatum, Melandrium triflorum, Potentitla hookeriana, P. chamissonis. Такое сообщество, конечно, не может быть названо степным.
Среди прочей растительности Чукотки степоиды, конечно, выделяются внешним обликом, особенно те, покрытие которых относительно велико, или те, в которых аспектируют злаки и осоки, внешний вид таких степоидов напоминает растительность пустырей в умеренных широтах с неравномерным покрытием и торчащими клочьями полусухих листьев злаков. Т. Г. Полозова (1981) в среднем течении р. Паляваам в степном флороценотическом комплексе насчитывает 32 вида: 1 — кустарничек, 2 — полукустарнички, 2 — многолетние монокарпические травы, 27 — поликарпические травы. Среди последних 12 видов стержнекорневых (в т. ч. 2 вида подушек), 5 — длиннокорневищных, 4 — плотнодерновинных, 4 — короткокорневищных, 1 — луковичных, 10 — надземноползучих. Преобладают растения с глубокой (до 50—60 см) стержневой корневой системой. Из-за доминирования травянистых мезоксерофильных форм степоиды и заслуживают выделения в качестве типа растительности, несмотря на то, что выделение более низких их категорий связано с едва ли разрешимыми проблемами фитоценологической классификации.
Флористические и ценотические различия между степоидами на лёссовидных суглинках на склонах высоких надпойменных террас и на щебнистых склонах гор не превышают различий между разными степоидами на одном и том же субстрате. В большинстве случаев эти различия значительны, т. е. степоиды мало походят один на другой флористически.
В бас. Анадыря на склонах гор встречаются сочетания степоидов с рощицами Betula platyphylla ssp . minutifolia и Alnus f ruticosa, напоминая внешним обликом аналог плейстоценового комплекса И. М. Крашенинникова (1939). Из полу- и ксерофильных кустарников в этих степоидах обычны Juniperus sibirica, Rosa acicularis и Rubus sachaiinensis, из полукустарничков — Dracocephalum palmatum, Thymus serpyllum, Artemisia kruhsiana, из суккулентов — Sedum cyaneum, Rhodiola rosea, из травянистых мезо ксерофите в — Phlojodicarpus villosus, Pulsatilla nuttalliana ssp. multifida, Veronica incana, Galium verum, Erysimum cheiranthoides, Artemisia tanacetifolia, Draba cinerea, Potentilla anadyrensis, Dianthus repens, Carex pediformis, C. obtusata и др. Флористическая насыщенность степоидов в значительной мере зависит от наличия на склоне выходов коренных пород, затрудняющих движение поверхностного субстрата вниз по склону. Флористический состав собственно чукотских степоидов включает Aster alpinus, Helictotrichon krylovii, Oxytropis campestris, O. adamsiana ssp. vassilczenkoi var. substepposa, O. middendorffii, Potentilla arenosa, Selaginella sibirica, Thymus serpyllum, Silene repens, Arenaria capillaris, Festuca ienensis, Eritrichium sericeum, Polygonum laxmannii, Bupleurum triradiatum, Cnidium cnidiifolium, Pulsatilla nuttalliana ssp. muIttfida, Lychnis sibirica ssp. samojedorum, Carex supina ssp. spaniocarpa, C. obtusata, C. duriuscula, Calamagrostis purpurascens, Smelowskia calycina ssp. integrifolia и др.
Было отмечено, что в среднем течении р. Амгуэмы (близ моста) существует флористическое ядро из константных и верных видов криофитностепных сообществ (Слинченкова, 1984). Некоторые виды ядра наблюдались нами в 1972 и в 1987 гг. не во всех этих сообществах (напр. Thymus serpyllum, Artemisia arctisibirica и др.) и, наоборот, некоторые наблюдались в горных тундрах, т. е. не были верными. В Зап. Гренландии степоиды или «субарктическая степь», по Бёхеру, возможно достигают 68° с. ш. ( Bocher, 1949). На Чукотке они распространены на север еще дальше. Степоиды есть, например, в окрестностях г. Певек. По данным Б. А. Юрцева (1974), степоиды (называемые им тундро-степными сообществами) имеются и на о-ве Врангеля, однако В. В. Петровский (1967) в своем очерке о растительности этого острова называл эту растительность (с доминированием Carex obtusata и С. rupestris) осочково-разнотравной тундрой.
В низовьях Колымы «реликтовые тундро-степные группировки» с доминированием Carex spaniocarpa описаны на 10 участках на сухих песчаных бровках озерных котловин и на вершинах гидролакколитов, но больше на Роговатской сопке, где 8 участков имеют общую площадь 20,5 кв. м (Андреев, Галактионова, 1981). Стало быть, средняя площадь одного участка немного менее 3 кв. м. Не удивительно поэтому, что только названная осока является стабильным видом на этих участках, а остальной набор видов сильно варьирует.
Степной тип
Ранее мы называли растительность этого типа каменисто-степной (Кожевников, 1974, 1976, 1979), но теперь, по изложенным выше мотивам, предпочитаем называть ее просто степной. Этот тип отличается от предыдущего только сомкнутостью растительности, флористический состав которой не обладает спецификой по сравнению со степоидами. Те отличия, которые могут обнаружиться между анадырскими и чукотскими степоидами, обусловлены пределами географического распространения видов, а не их фитоценологическими предпочтениями. Среди доминантов степного типа нужно назвать Carex supina ssp . spaniocarpa, С. pediformis, C . duriuscula, Festuca Ienensis, Calamagrostis purpurascens, Elymus confusus. Можно отметить, что степные участки, как правило, бедны флористически, но зато разнообразны по строению, в связи с чем возникает сомнение — настолько ли сильно ценотическое давление в этих сообществах, чтобы их отличать от степоидов по этому признаку. Однако даже в Туве (в Хемчикской котловине) степная растительность наиболее разнообразна по структуре и бедна флористически (всего 10—25 видов) (Намзалов, 1978). Среди них имеются виды, общие с анадырско-чукотским регионoм : Carex duriuscula, C . pediformis, Veronica incana, Galium verum, Artemisia frigida.
Степные участки заметны издалека благодаря соломенной окраске. На некоторых из них безусловно существует ценобиоз. Степной тип находится в таком же отношении к степоидному, как лесной к лесотундровому. Поэтому, если не признавать самостоятельности первых двух, то и вторые нужно рассматривать совместно. Степные участки более характерны для Анадырского края, чем для Чукотки, где они крайне редки. Для нас сейчас важно подчеркнуть географический параллелизм в проявлении лесного и степного типов растительности, а также их параллельную трансформацию в другие типы вследствие утраты или ослабления ценобиоза, а также их заметную современную деградацию в связи с происходящим изменением климита (см. ниже). Степные сообщества Западной Гренландии имеют рН почв 6,7—8,9 (Bocher, 1949). По Чукотке таких данных пока нет и можно лишь догадываться, что многие степные участки имеют кислую среду из-за накопления органических остатков, а другие — нейтральную из-за выщелачивания. Косвенным свидетельством кислой реакции субстрата на степных участках является произрастание под этими участками Comarum palustre, который населяет озерки, имеющие низкие (кислые) значения рН (Bocher, 1949).
На Колымском нагорье М. П. Данилов расценивает совокупность видов степных склонов и известняков как один флористический комплекс, так как «их объединяет один эдафический фактор — высокая карбонатность субстрата» (Данилов, 1978. С. 39). Автор, однако, не провел флорогенетически и анализ своего материала, ограничившись ссылками на более, чем сомнительные источники, поэтому причины совместного произрастания степняков и кальцефитов не раскрыты. По-видимому, это обусловлено физическими свойствами известняков, благоприятными для существования степняков, а вовсе не «высокой карбонатностью субстрата», выступающей в данном случае как побочный фактор. Длительный прогрев субстрата, т. е. накопление значительной суммы температур, является важнейшим фактором развития и существования степной растительности. Указано, что в зональных степях и пустынях суточная амплитуда температуры у поверхности почвы составляет 15—20° (30°) (Хромов, 1968). Отрывочные замеры в чукотских степоидах говорят о том, что суточные амплитуды на поверхности их почв могут достигать не меньших, а иногда и больших значений (до 35°).
Горнотундровый тип
Растительность этого типа обычно называют щебнистой тундрой, поскольку ее покрытие часто очень незначительно, и основной фон земной поверхности создает щебень, а не растительность. Однако в щебнистых тундрах горных склонов наблюдаются плавные переходы одних сообществ и группировок в другие в связи с изменением рельефа, обычно крутизны. Кустарничковая горная тундра при увеличении угла наклона переходит в редко-кустарничковую тундру, так как те же самые кустарнички имеют здесь меньшее обилие. Существуют и резкие изменения растительности, но почти всегда они связаны с изменением микро- или даже нанорельефа. К этому типу относятся также лишайниковые варианты разреженной кустарничковой растительности. Судя по тому, что флористически (по высшим растениям) и структурально участки с лишайниковым ковром и без него при прочих равных не различаются, лишайниковая синузия не оказывает влияния на многие тундровые виды, хотя лишайники, как известно, подкисляют субстрат.
Тип составляют формации тундр дриадовых, вороничных, диапенсиевых, луазелериевых, кобрезиевых, лишайниково-кассиопейных, осочковых (с Carex rupestris), а также редкотравно-пустошные, смешанно-кустарничковые и т. п. Дриадовые тундры с доминированием Dryas octopetala ssp. viscida распространены в основном на кислых и средних породах. На основных и щелочных породах доминируют D. octopetala s. str. и D. integrifolia. Горные тундры с господством D. octopetala ssp. punctata (D. punctata) Б. А. Юрцев (1966) называет «наиболее ксеротермными разностями горных тундр», что не соответствует действительности даже в отдельных случаях. Они не являются более ксеротермными, чем другие горные тундры, перечисленные выше. Горнотундровый тип растительности лесных и лесотундровых районов Анадыря весьма отличается флористически от районов Чукотки отсутствием многих арктоальпийских видов и родов Minuartia, Draba, Saxifraga. Позитивное отличие анадырских горных тундр от чукотских значительно скромнее. Оно заключается только в наличии Sedum cyaneum. На Чукотском полуострове имеются эдификаторы горных тундр Rhododendron camtschaticum, Carex nesophila, которые в континентальных районах находятся в реликтовом состоянии или отсутствуют.
Для горнотундрового типа характерна мозаичность растительности, обусловленная либо случайным распределением растений, либо нанорельефом, обусловливающим некоторые различия в среде обитания: почвенных и температурно-влажностных условий. Последнее сказывается прежде всего на покрытии растительности. Так, на поверхности высокой надпойменной террасы р. Янрамавеем с куртинной дриадово-лишайниковой тундрой покрытие высших растений на взлобке составляло 20%, а в ложбинке — 60%. Соответствующие температуры в почве были 13° и 10°, а на ее поверхности 14,2° и 13°. Флористический состав также различался (общие виды заключены в скобки): Artemisia furcata, Senecio tundricoia, Potentilla elegans (Dryas octopetala ssp. viscida, Carex nesophila, Oxytropis czukotica, Hierochloe alpina), Salix sphenophylla, Diapensia obovata, Rhododendron parvifolium. Хотя флористическое различие кажется значительным, так как общих видов 4, а не общих 6, оба набора относятся к одной и той же растительности. Подобных примеров можно привести много. Флористический состав горнотундровых сообществ и группировок насыщен аркто-альпийскими видами. Характерно обилие камнеломок и крупок.
На севере Чукотки существует особый вариант горных тундр. Эти тундры характеризуются очень малым покрытием растительности, которая почти не возвышается над поверхностью галечного субстрата. Характерным местоположением является очень выровненная поверхность галечной толщи, выстилающей приморские равнины, т. е. это — плакорная растительность. Доминирует Dryas octopetala ssp. viscida. Кроме того, здесь велико обилие лишайников, особенно Alectoria nigricans, сообщающей участкам черноватый цвет. Эти участки отличаются также зачернением гальки, покрывающей их поверхность. Очевидно, это обусловлено выцветами марганца, что характерно для районов высокой Арктики с ее малым количеством осадков и для пустынь и полупустынь (Горбацкий, 1964). Действительно, почернение камней здесь очень напоминает так называемый пустынный загар.
Луговинно-нивальный тип
Этот тип включает два подтипа: луговинный и нивальный. Первый представляет преимущественно сомкнутую мезоморфную растительность, с кустарничками, среди которых особенно характерны Salix chamissonis, Pentaphylloides fruticosa, Spiraea stevenii. Травянистые эдификаторы — Festuca altaica и Сагех podocarpa. Среди обязательных компонентов нужно назвать Artemisia arctica, A. tilesii, Polemonium acutiflorum, Aconitum delphinifolium, Anemone sibirica, Poa malacantha, Trisetum spicatum, Saxifraga nelsoniana, Potentilla hyparctica, Valeriana capitata и др.
|

Луга и ивняки в пойме в окрестностях п. Лорино на Чукотском полуострове. Пар над рекой - следствие выхода на поверхность горячих вод (горячие источники). Фото А.А. Галанина
|
Согласно Л. Г. Раменскому (1925), травяные ценозы настолько подвижны и пластичны, что быстро достигают практического равновесия, т. е. приблизительного количественного равновесия основных массовых видов. Это суждение вполне применимо к луговинной растительности Чукотки, поскольку описания ее из даже разных районов обычно очень сходны. Особый вариант луговинной растительности представлен на сусликовинах (Тихомиров, 1960).
Нивальный подтип представляет растительность из хионофильных видов, сформировавшуюся под влиянием талых вод от наледей и снежников, стаивающих не раньше середины июля или перелетков. Мощный снеговой покров, по Б. А. Тихомирову (1945), имеет огромное экологическое значение. Он предохраняет растения от воздействия низких температур, создает равномерное непрерывное проточное увлажнение, понижает уровень вечной мерзлоты, сдвигает фенофазы растений, создает неблагоприятные условия для развития споровых растений, а также деревьев и кустарников, при стаивании образует наилок.
Характерные виды этой растительности Сагех tripartite, Koenigia islandica, Ranunculus pygmaeus, R. nivalis, Stellaria irrigua, Poa paucispicula, Salix polaris, Saxifraga hyperborea, S. nelsoniana ssp. porsildiana, Rumex arcticus, Phippsia algida. |
В зависимости от продолжительности стаивания снежников, которые образуются или подновляются на одном и том же месте каждый год, четко выделяются три варианта нивальной растительности: кассиопейная (с доминированием Cassiope tetragona) - снежники стаивают в июле; ковровые ивковые (с доминированием Salix polaris) — снежники стаивают к августу; редкотравные подснежниковые группировки —снежники стаивают к сентябрю или не дотаивают. Эти группировки обычно находятся на переувлажненном илистом субстрате, являющемся продуктом нивации. На нем обычны специфические мелкие мхи, придающие отдельным участкам изумрудный цвет. В. Д. Александрова (1977) выделила эти группировки (судя по ее описанию) в особый тип растительности — высокоарктические минеральные болота.
В луговинной растительности, согласно Б. А. Тихомирову (1945), отсутствуют ценотические отношения, и условия существования этой растительности регулируются физическими факторами — снеговым покровом и процессами, вытекающими из продолжительности его лежания, интенсивности таяния. В связи с этим структура нивальной растительности сильно зависит от местоположения снежников, поэтому в одних случаях главенство получают виды, которые в других случаях играют второстепенную роль. Разнообразны и переходы нивальной растительности в луговинную, которые прослеживаются при удалении от края снежника. Лучше всего эти переходы видны на склонах надпойменных террас в местах, где у подножия снеготаяние задержано. В таких случаях у подножия склона существуют типичные нивальные сообщества, а в верхней его части — типичные луговинные сообщества; посередине наблюдается плавный переход.
В качестве примера приведем один такой случай из верховьев р. Гытхытвыэоуваам. У подножия склона (25°) в долину горного ручья описана бугорковатая нивальная луговина с фоном Salix polaris, которая включает Alopecurus alpinus, Draba juvenilis, Сагех podocarpa, С. tripartite, Eritrichium villosum, Artemisia tilesii, Poa malacantha, Polemonium acutiflorum, Oxyria digyna, Artemisia arctica, Luzula multiflora, Saxifraga nelsoniana ssp. porsildiana, Ranunuculus nivalis, Saxifraga hyperborea, Acomastylis rossii, Potentilla hyparctica, Taraxacum lateritium, Allium schoenoprasum. В верхней части склона расположена мезоморфная луговина с фоном Salix shamissonis, включающая Pentaphylloides fruticosa, Pedicularis capitata, P. oederi, Polygonum tripterocarpum, P. viviparurn, Potentilla hyparctica, Dryas octopetala ssp. viscida, Festuca altaica, Vaccinium uliginosum ssp. microphyllum, Saxifraga nelsoniana, Antennaria dioica, A. monocephala, Carex nesophila, Aconitum delphinifolium, Trisetum spicatum, Hedysarum hedysaroides ssp. arcticum, Cerastium alpinum ssp. jenisejense, Eritrichium villosum.
В местах выхода на поверхность грунтовых вод от подтаивания мерзлоты часто сформированы сообщества, являющиеся промежуточными между типично нивальными и типично луговинными. Это обстоятельство, наряду с тем, что оба подтипа часто связаны пространственно, склоняет нас рассматривать луговинно-нивальную растительность в качестве одного типа, а не двух, как мы считали раньше (Кожевников, 197бж, 1978в). К этому надо добавить, что оба подтипа имеют на Чукотке сходное (параллельное) географическое распространение, тяготея к океаническим районам. Аналогичная ситуация наблюдается в Гренландии, где у побережий имеются снежники и травяные луговины, но во внутренних континентальных районах нет ни того, ни другого (Bocher 1949). Б. А. Тихомиров (1945) расценивал нивальные луговины (луга, по его терминологии) как климатогенные, отличая их по обусловленности от экогенных приручьевых и приморских луговин.
Высокоарктический тип
Под этим типом мы понимаем сильно разреженную травяную растительность близ северного побережья Чукотки. Доминируют в ней злаки. В Ванкаремской низменности в этой растительности наиболее представлена Arctagrostis arundinacea, часто с содоминированием Carex bigelowii ssp. lugens. Местоположение этой растительности — верхние части плоских увалов. По сравнению с подобными тундрами на о-ве Врангеля материковые тундры высокоарктического типа довольно богаты флористически; часто в них обильны кустарнички Dryas octopetala ssp. viscida, Salix sphenophylla, S . phlebophylla, Diapensia lapponica ssp. obovata, но гипарктические кустарнички отсутствуют. Эти тундры можно рассматривать как редуцированный вариант луговинной растительности, однако они обладают специфической географией, т. е. находятся в таком же отношении к луговинно-нивальному типу, как лесотундровый к лесному или степоидный к степному. Подобная, но еще более редуцированная растительность арктических островов была выделена в качестве полярнопустынного типа (Александрова, 1971, 1977; Короткевич, 1972).
Типы растительного покрова
Теперь обратимся к типам растительного покрова, поскольку именно они являются основой ботанико-географического деления земной поверхности. Это отчетливо выражено А. Н. Лукичевой (1967) в «Физико-географическом атласе мира», где она различала «бореальный тип растительности», включающий не только господствующие формации, но и генетически связанные с ними, например, болота и луга внутри ареала бореальных лесов. В данном случае понятие «тип растительности» был смешан с понятием «тип растительного покрова».
Говоря о распределении типов растительности Анадырского края, В. Н. Васильев (1956) показывает в действительности топологические особенности растительного покрова (7 типов) по преобладающим формациям. Например, долинная растительность, по Васильеву, может рассматриваться в общих чертах в пределах трех типов: 1) растительность верховий рек, 2) растительность средних течений, 3) раститель ность низовий. Здесь опять прослеживается смешение понятий «тип растительности» и «тип растительного покрова».
Оба эти понятия, разумеется, взаимосвязаны, и характер этой связи определяет отнесение территории к той или иной зональной и провинциальной категории. Взаимосвязь этих понятий по принципу обратной связи позволяет конкретизировать различия некоторых типов растительности и отнесение определенных участков к тому или иному типу. В этом отношении особенно нагляден пример района среднего течения р. Майн. Л. Н. Тюлина (1936) писала, что на Майне расположена собственно лесотундра, а в центре Верхне-Анадырского района встречаются уже первые островки настоящего леса. Однако то, что на Майне принимается за тундровую составляющую растительности, на самом деле является болотом. Настоящие тундры в этом районе отсутствуют. Нет их и в южной половине марковской впадины. Обнаружено также, что и флора, и фауна обоих названных районов состоит преимущественно из лесных северобореальных представителей; тундровые виды представлены как большая редкость или отсутствуют. То, что А. П. Васьковский (1958) называл «лесом на тундре», в действительности является лесом на болоте, что очень характерно для северной тайги.
Хотя Л. Н. Тюлина (1936) пришла к выводу, что полоса криволесья и сухостоя у предела лесов в Анадырском крае не выражена, она же отмечает, что «граница лесной зоны здесь выражена не сплошными лесными массивами, а лиственничным редколесьем (с ярусом кедровника) по склонам сопок, которые еще нельзя назвать лесом, а также и плохо сомкнутыми (куртинного и ленточного характера) древостоями лиственницы, распространенными по надпойменным террасам» (Тюлина, 1936. С. 44). Нам, однако, удалось найти на северной окраине майнского таежного массива крупный, высохший на корню лиственничник и такой же березняк. Вероятно, имеются и другие сухостойные леса, еще не обнаруженные. Лесные формации бассейна среднего Анадыря, к которым мы относим и заросли кедрового стланика, сочетаются в ландшафтах с лесными травяными болотами, лугами, кустарниковыми лугами, лишайниковыми парковыми зарослями кустарников и зарослями из бореальных высокорослых ив вдоль рек.
Район среднего течения Майна существенно отличается от района Марковской впадины широким распространением лиственничников, которые нацело отсутствуют в марковской впадине. Но это не единственное отличие. В среднем течении Майна распространены ольховые леса из Alnus hirsuta. В Марковском районе этот вид совсем не представлен. К этим различиям добавляются многочисленные флористические различия. Представление В. Н. Васильева (1956) о том, что тополевники сменяют чозенники в ходе исторического развития растительности оказалось неверным (Кожевников, 1981). Напротив, в окрестностях Марково наблюдается деградация тополевников, которые к тому же и очень редки в данной местности (скорее это тополевые колки) и имеют очень разреженный древостой тополя. Еще Л. Н. Тюлина (1936) отметила на р. Марковке засыхающий тополевник, что упоминает и В. Н. Васильев (1956. С. 155). До времени нашей работы в Маркове в 1974 г. этот тополевник еще не засох до конца, хотя попрежнему производит впечатление засыхающего. Однако выглядит он теперь более разреженно, чем на снимке Л. Н. Тюлиной (1936, с. 41). Кроме того, он уже не столь просветлен, в нем развилась мощная ивовая поросль. Тем не менее это, пожалуй, один из единичных в окрестностях Маркова тополевников, редкостойный как и все прочие. Вместе с тем, в окрестностях Маркова развиты великолепные чозенники до 30 м высотой. Обратные отношения этих деревьев наблюдаются на Майне. Здесь развиты великолепные тополевники, а чозения всегда отмечается лишь в виде незначительной примеси. При этом нет никаких оснований думать, что все чозенники на Майне вытеснены тополевниками. В. Н. Васильев (1956. с. 154 ) упоминает о тополевнике в долине р. Ваеги (приток Майна) с полнотой насаждения 0,7. При этом насаждения тополя — 0,9, а чозении — 0,1. (Тут, вероятно, опечатка, т. к. при общей полноте 0,7 насаждения тополя не могут иметь полноту 0,9. Вероятно, должно быть 0,6. На месте отмирающих тополевников, по мнению Тюлиной (1936), развиваются в Марковском районе парковые тальники). Такие отношения весьма характерны для всего Майна, и часто чозения играет еще меньшую роль.
Лесотундровый тип растительного покрова характеризуется сочетанием формаций лесотундрового типа растительности и субарктических болот и тундр, причем среди последних есть много промежуточных вариантов. В 1975 г. нам пришлось убедиться в том, что лесотундра делится на две полосы флористически. В южной полосе, примыкающей к северной тайге, отсутствует большое количество тундровых видов. В северной полосе они появляются в изобилии (Кожевников, 19776). Обусловлено это близостью гор, к которым идет постепенное повышение местности, в связи с чем появляются пятнистые тундры, представляющие, по В. Б. Сочаве (1930), растительные сообщества, среди которых равномерно, в виде более-менее округлых пятен, выходит на поверхность подпочва — горизонт С. В южной полосе анадырской лесотундры на разных местообитаниях еще встречаются крупные одиночные березы, на болотах растет стланик, в укрытых местоположениях вдоль проток встречаются ивовые лески из Salix udensis и одиночные деревья Alnus hirsuta. В северной полосе вне пойм растет лишь кедровый стланик, который избегает здесь селиться на заболоченных шлейфах гор. Все изменения в растительном покрове объяснимы физико-географическими условиями, а не конкуренцией разной растительности.
По мнению М. Фернальда (Fernald, 1925), представители арктической флоры из-за своей молодости дали мало локальных эндемиков и обладают повышенной жизнеспособностью по сравнению с представителями более древней бореальной флоры. Поэтому арктические виды, как считал Фернальд, способны вытеснять бореальные виды там, где они находятся в соприкосновении. Аналогичную мысль высказывал Б. А. Тихомиров в 1946 г. «...Холодная волна послеледникового времени оживила арктические элементы и заставила многие теплолюбивые растения отступить на юг». Эта точка зрения во многих своих пунктах весьма спорна. Прежде всего совершенно неправильно воззрение, что арктические виды мало дифференцированы на различные локальные расы. Скорее даже наоборот, их расовая дифференциация, именно из-за относительной молодости, превосходит таковую бореальных видов. Взаимоотношения арктических н бореальных видов в районах их стыковки могут быть самыми разными, но арктические виды не могут вытеснять бореальные, если этому не способствует изменение среды. Об этом свидетельствуют палеогеографические факты. Как только наступало потепление климата, бореальные виды быстро заселяли Арктику и вытесняли арктические виды, М. Порсилд (Porsild, 1920) отметил на о-ве Диско, что отсутствие арктических растений на низменных равнинах вряд ли обусловлено климатически, скорее это — результат конкуренции.
Вертикальная поясность в анадырской лесотундре — кедровостланиково-гольцовая. В. Н. Васильев (1956) относит полосу кедрового стланика к подзоне кустарниковой тундры, отличая ее от полосы ольховника. Полосы эти прекрасно прослеживаются в природе. А. П. Васьковский (1958) даже предложил для них особые названия: кедро-тундра и ольхо-тундра. Однако на основании ботанико-географических и, даже шире, биогеографических связей и отношений и специфической биологии кедрового стланика его заросли следует рассматривать как лесной тип растительности, а чередование его зарослей с кустарничковьми формациями — как лесотундровый тип растительного покрова. В этом типе еще велика роль устойчивых ценотических систем с высоким биотическим давлением и, следовательно, стабильными флористическим составом и структурой. Эти ценотические системы, относящиеся к разным типам растительности, выходят в лесотундру из северной тайги, постепенно уменьшаясь в численности и типов и представительства внутри отдельных типов. Вместе с исчезновением при движении к северу таежных ценосистем, исчезает сразу большое количество видов, связанных с этими системами. В данном случае мы имеем хорошее соответствие с представлениями С. М. Разумовского (1969).
Б. Н. Норин (1967, 1979) доказал независимость ярусов в лесотундре, установив, что в пределах яруса растения образуют вполне закономерные объединения, которые могут быть классифицированы. Эти микрогруппировки разных ярусов в значительной мере самостоятельны и независимы друг от друга. Это представление геоботаника находит отражение в ботанической географии анадырско-чукотского региона, где в некоторых многоярусных ценосистемах при движении к северу происходит выпадение верхних ярусов. Уже в лесотундре некоторые кустарниковые сообщества представляют не что иное, как растительность подзоны северной тайги, утратившей древесный ярус. При движении далее к северу утрачивается и кустарниковый ярус; остается лишь наземный кустарничково-моховой ярус. В этой связи В. С. Говорухин (1951) писал, что нередко целые синузии (например, целые ярусы — травяной, моховой и др.) образуются, складываются в одной растительной формации, но затем путем миграции внедряются и в другие формации. Имеются, однако, основания полагать, что наличие одних и тех же синузий в разных формациях и типах растительности складывается не только в результате миграции, но и в результате редукции других синузий (особенно верхних ярусов) при изменении климата.
Северная граница лесотундры до низовьев Колымы на западе является сухопутной границей Чукотки как природной области. Эту границу мы проводим по пределам распространения Pinus pumila, являющегося хорошим индикатором растительного покрова, в котором преобладают бореальные формации. Только в районе Майна-Пыльгина кедровник выходит к морскому побережью (Васильев, 1956).
Мы совершенно согласны с Л. Хамет-Ахти (1976), что к бореальному типу растительного покрова следует относить не только лесные формации, но и определенные луговые, болотные и др. Таким образом, лесотундровый тип ландшафта может включать исключительно безлесные фации, но, как правило, в нем имеются хотя бы одиночные деревья. Северная граница лесотундры в таком понимании является не только ботанико-географической. Поскольку в лесотундре обитают таежные представители фауны, то и зоологи проводят ее северную границу по пределам распространения кедрового стланика (Портенко, 1939; Кищинский, 1970). Наши собственные наблюдения за птицами на Чукотке и в Анадырском крае (Кожевников, 1977а, б; 1982) дают дополнительное представление о том, что состав орнитофауны в анадырской лесотундре ближе к составу северной тайги этого региона, чем к подзоне кустарниковых тундр южной Чукотки, но тундровые виды здесь встречаются, причем приурочены они именно к тундровым участкам. По данным В. А. Кузякина (1967), европейская лесотундра характеризуется прежде всего специфическим комплексом птиц, распространенных в целом как севернее, так и южнее лесотундры.
Таким образом, лесотундра представляет собой переходную полосу между таежной и тундровой зонами, но в силу своей обширности и достаточно выраженной специфики ее также наделяют рангом зоны. Исходя из сказанного, мы не можем согласиться с точкой зрения В. Д. Александровой (1977) об отнесении Корякского нагорья и Анадырско-Пенжинской территории к Арктике. И то и другое должно относиться к зоне лесотундры, как это и было принято авторами «Геоботанического районирования СССР» (1947) или, во всяком случае, к бореальной области (Юриев, 1967). К северу от лесотундры типы растительного покрова сильно подчинены влиянию географического окружения местности. Так, на Анадырском плато, подвергающемся воздействию ветров с Восточно-Сибирского моря, на днищах долин кустарниковая растительность очень угнетена и практически отсутствует. Но по соседству, в Амгуэмо-Куветском горном узле, в межгорных впадинах формируется собственный устойчивый физико-географический процесс континентального типа. Поэтому в межгорных впадинах Амгуэмо-Куветского узла развита кустарниковая растительность, встречаются чозениевые рощи. Кустарниковая растительность в некоторых местах поднимается на склоны гор до 200 м относительной высоты. На шлейфах гор развиваются мощные моховые покровы, в которых велика роль сфагновых мхов. Встре чаются степоиды и, как большая редкость, степные сообщества. Ряд континентальных видов отличается высокой активностью, полностью отсутствуя на Чукотском полуострове с его океаническим климатом, например, Salix tschuktschorum. Такие виды, как багульник, березка, голубика, имеют кроме кустарничковой биоморфы и низкокустарниковую и в совокупности или по отдельности образуют густые заросли. Как отмечено В. Н. Андреевым (1966), форма роста березки является указателем зонального характера растительного покрова: ортотропный рост — южнотундровая растительность, плагиотропный рост — северотундровая растительность. Оба типа роста обусловлены как внешними факторами (давление среды), так и внутренними (биологическими) (Серебряков, 1962).
|

Чукотский полуостров. Оленье стадо в типичной тундре. Фото А.А. Галанина |
Таким образом, растительность межгорных впадин Амгуэмо-Куветского узла является южнотундровой, кустарниковой. Здесь сочетаются кустарниково-гольцовый и тундрово-гольцовый типы вертикальной поясности. Орнитофауна включает целый ряд бореальных видов, населяющих разные стации, особенно островные леса.
Согласно Л. И. Добринскому (1967), в островном Хадытинском лесу в тундре Приобского Севера существует комплекс гнездящихся видов птиц, составленный типично тундровыми и типично лесными представителями орнитофауны. При этом некоторые виды, как утверждает автор, вошли в этот комплекс лишь в последнее десятилетие. И. В. Игнатенко (1967) показал для восточноевропейской лесотундры большое сходство болотистых почв в подзонах лесотундры и кустарниковых тундр, что, по его мнению, подтверждает справедливость отнесения кустарниковых тундр к лесотундре. Очевидно, следует изучить в том же плане и болотистые почвы подзоны типичных тундр, чтобы подчеркнуть их зональные отличия от аналогичных почв лесотундры. Этот же автор получил убедительные зональные отличия почв голых тундровых пятен в подзонах типичных и кустарниковых тундр (Игнатенко и др., 1967). Таким образом, кустарниковые тундры ближе к лесотундровому типу растительного покрова, чем к типично тундровому, что с геоботанических позиции показано также Б. Н. Нориным (1979).
|
По мнению В. В. Крючкова (1967), к северной подзоне лесотундры относятся кустарниковые и кустарничковые формации и лесные островки. Если придерживаться этой точки зрения, то северная граница северной подзоны лесотундры должна проводиться от верховий Анадыря через среднее течение р. Белой, вдоль р. Осиновой (притока р. Белой) вдоль северных отрогов Амгуэмо-Куветского массива до р. Амгуэмы, затем от Амгуэмы на юг и выходить к верховьям зал. Креста. Однако кустарниковая и, тем более кустарничковая, растительность межгорных впадин Амгуэмо-Куветского массива и Анадырской низменности не относится к лесотундровому типу растительного покрова. Генетически и формационно эта растительность близка к субальпийской растительности северной Охотии и Камчатки, впрочем, как и кедровниковая. Хотя в этих районах встречаются чозениевые рощи и небольшие березнячки, которые занимают исключительно благоприятные местообитания и редки, чтобы при обозначении типа растительного покрова их учитывать и говорить о лесотундре. Все эти осколки лесного типа растительности носят реликтовый характер. Они имеют вес лишь при характеристике окружающего их растительного покрова, но в зональном отношении только усиливают отнесение очерченного района к подзоне кустарниковых тундр. Б. Н. Городков (1935) называл острова леса в тундре «тундровыми лесами», причисляя их, однако, к лесному типу растительности.
Кустарниковый тип растительности встречается и в подзоне типичных тундр, где он приурочен к поймам и склонам в речные долины. Он отсутствует только близ морских побережий, северных низменностей. Уже в подзоне кустарниковых тундр, а в еще большей мере в подзоне типичных тундр мы не находим существенных сгущений границ ареалов видов со сходным географическим распространением. Разумовский (1969) отметил, что «наибольшее число примеров несовпадения границ показывает группа видов, не входящих в ценотические системы и существующих в специфических условиях отсутствия биотического давления». Исходя только из этого, можно судить о том, что уже в подзоне кустарниковых тундр биогенная компонента среды обитания значительно ослаблена, хотя в ряде случаев мы не можем не считаться с ней.
Типы растительного покрова в типично тундровых ландшафтах включают более дробные сочетания участков, чем в ландшафтах кустарниковых тундр, а в этих более дробные, чем в лесотундровых ландшафтах. Таким образом, с деградацией ярусов возрастает комплексность и мозаичность растительности.
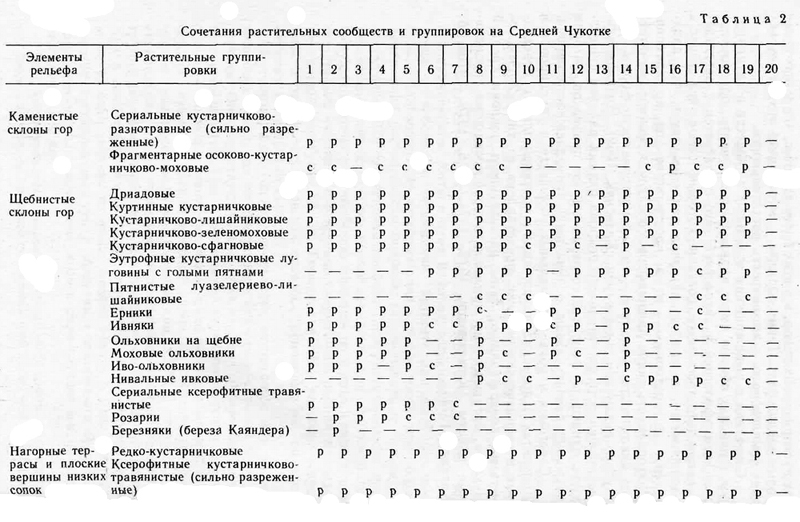


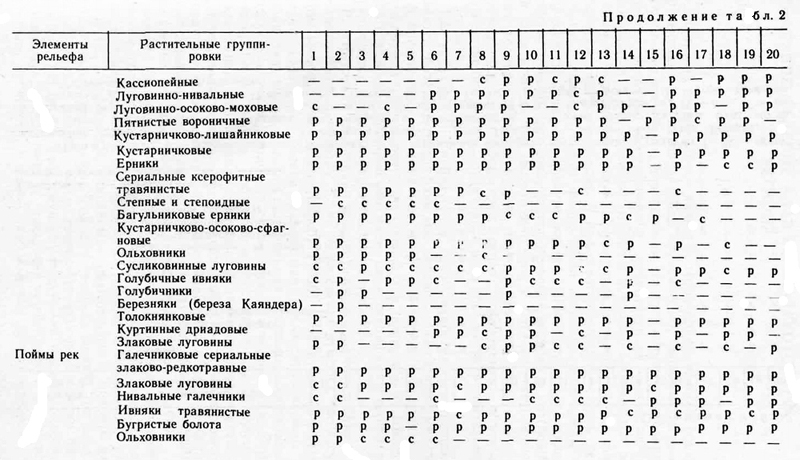

Сочетания и выраженность отдельных сообществ и группировок в районах обычно имеет индивидуальные черты, обусловленные геоморфологией и литологией. Это согласуется с представлениями ландшафтоведов, согласно которым имеющийся сейчас в распоряжении географов материал свидетельствует, что ландшафты не повторимы ни в пространстве, ни во времени. Повторяемость типов ландшафтов, создающая ложное впечатление цикличности развития, — только повторяемость формы, при глубоко различном содержании (Калесник, 1952). Индивидуальные особенности растительного покрова разных районов существуют, однако, на фоне той совокупности его признаков, которая позволяет сделать вывод о зональной принадлежности района. Сочетания и роль растительных сообществ и группировок в 20 пунктах Чукотки, расположенных в континентальных, переходных и океанических районах, представлена на табл. 2. Эти сочетания конкретизируют типы растительного покрова средней Чукотки, ботаническому описанию которой посвящены специальные работы (Кожевников, 1973, 1974, 1976, 1978, 1979а, б; Катенин, 1974).
Читать следующую главу