|

Мая Тимофеевна Мазуренко и Андрей Павлович Хохряков |
Электронное издание этой книги подготовлено М.Т. Мазуренко на основании бумажного варианта, изданного в 1977 г. Сканирование текста книги и рисунков выполнено в рамках Аналитической ведомственной целевой программы "Развитие научного потенциала высшей школы (2009-2010 годы) по проекту "Развитие Ботанического сада Южного федерального университета в качестве ресурсного, учебного, информационного и культурного центра на юге России".
Редактор электронного издания А.В. Галанин
Приводится описание структуры и циклов развития побегов и побеговых систем более 100 видов дикорастущих или экзотических кустарников и кустарничков из различных ландшафтно-климатических зон СССР главным образом по материалам, собранным авторами в течение 1969-1975 гг. Большое внимание уделяется изменчивости побеговых систем кустарников в зависимости от условий существования. Для 18 видов дается подробное описание фаз развития от прорастания семени до отмирания. Приводится краткая классификация побегов и побеговых систем всех древесных растений. Жизненная форма кустарника (кустарничка, полукустарника) рассматривается как функция циклов развития; побеговых систем, определяющая длительность той или иной фазы онтогенеза и зависящая от изменения условий существования.
Монография рассчитана на специалистов-ботаников, студентов биологических вузов, садоводов разного профиля, широкий круг биологов, интересующихся проблемами метамерии и системного подхода к биологическим явлениям.
Табл. 3, ил. 50, список лит. 66 назв.
Ответственный редактор А. Г. КРЫЛОВ |
 |
Введение
Кустарники представляют собой одну из обычных жизненных форм семенных растений, широко распространенную почти по всей поверхности нашей планеты, исключая самые пустынные области арктических широт, аридных пространств и альпийских высокогорий. На обширных территориях они составляют основу растительного покрова, как, например, в средиземноморских маквисах и шибляке, австралийских срёбах, южноамериканских чаппаралях, кустарниковых и кустарничковых тундрах. Очень заметную роль они играют в растительном покрове саванн и степей.
Интерес к кустарникам вызывается также и тем, что многие из них представляют собой большую хозяйственную ценность. В первую очередь, это касается декоративных, широко используемых в озеленении кустарников, таких как многочисленные сорта сирени, чубушников, акаций, спирей, жимолостей. Не менее широко представлены в культуре и ягодные кустарники, такие как вишня, малина, смородина, крыжовник. Все более интенсивно используются и заросли дикорастущих кустарников. Ягоды многих из них служат витаминным сырьем, роль которого особенно велика на северо-востоке Сибири. Наиболее широко в качестве источника витаминов известен шиповник. В Сибири же, особенно Восточной, па первый план в этом отношении выступают смородины и жимолость съедобная.
Значительный интерес представляют собой кустарники и как жизненная форма, занимающая промежуточное положение между деревьями и травами. Именно у них возникают и развиваются те особенности структуры и морфогенеза, которые с течением эволюции привели к образованию многочисленных и разнообразных травянистых растений. |
С морфологической точки зрения кустарники — древесные растения, обладающие несколькими, обычно многими главными (основными) скелетными осями — стволами и стволиками, среди которых невозможно или очень трудно, да и то только на первых стадиях онтогенеза, выделить главный. Обычно при этом главные скелетные оси (стволики) более или менее быстро сменяют друг друга во времени таким образом, что онтогенез кустарника, его «большой жизненный цикл», складывается из большего или меньшего числа более коротких «онтогенозов» (жизненных циклов) его основных скелетных осей. Эти последние, в свою очередь состоят из единиц (метамеров) более низкого порядка — приростов, побегов и их систем, которые также имеют свои, еще более интенсивные циклы развития.
Цель настоящей работы - изучение структуры и морфогенеза жизненной формы кустарника. Основное внимание мы уделили метамерному строению и развитию побеговых систем, понимая под этим повторяемость некоторых характерных их структур во времени и пространстве, т. е. цикличность. Хотя изучению метамерии и отдельных сторон строения и развития различных жизненных форм в настоящее время придается большое значение, тем не менее вопрос о цикличности развития побеговыхсистем, соподчинении разных циклов и даже о самом количестве соподчиненных циклов в развитии тех или иных структур у растении в целом еще очень далек от разрешения. Свою работу мы рассматриваем поэтому только в порядке его постановки вовсе не претендуя на окончательное разрешение. Хотим лишь подчеркнуть, что большой жизненный цикл (онтогенез в целом) слагается из большего или меньшего числа более мелких циклов, и на примере кустарников это выступает очень наглядно. Однако прежде чем приступить к непосредственно интересующей нас проблеме циклов развития побеговых систем кустарников и их разнообразия, считаем необходимым дать обзор строения элементарных единиц, из которых складывается тело кустарников, побегам и их приростам.
Глава I
Разнообразие побегов и их систем у кустарников:
основные типы побегов
Типы побегов
Рост, развитие, поведение различных жизненных форм растений в разных экологических условиях зависят прежде всего от биологических особенностей и морфологии их побегов и образуемых ими систем. Отсюда вытекает необходимость рассмотреть прежде всего вопрос о разнообразии побегов и их свойств, в частности у кустарников. К побегам можно подходить с разных позиций. Очень хорошо изучены к настоящему времени морфология, т. е. форма, внешний вид побегов и их внутреннее строение. Достаточно полно известна к настоящему времени и биология побегов — способы их нарастания, циклы развития, длительность жизни и т. д, И наконец, только в начальной стадии находится разработка вопросов о побеге как основной единице метамерного тела растения. Это необходимо в целях выяснения гомологии частей различных жизненных форм или внутри одной понимаемой широка жизненной формы или даже отдельных побегов одного растения.
С морфологической точки зрения побег — это стебель с сидящими на нем листьями, их пазушными почками и иными придатками (волосками, чешуйками, колючками и т. д.). Но и простая соломина злаков, и ветвистый ствол дуба — одинаково стебли с сидящими на них листьями. Поэтому важно договориться о том, что считать побегом в качестве единицы метамерного тела растения, а что системой побегов. В настоящее время многие исследователи (Грудзинская, I960: Петров. 1963; Нухимовский, 1970; Гатцук, 1970, 1974а) единодушны в том мнении, что побег - это, собственно, простой неветвистый стебель, вырастающий из почки зародыша или любой другой, находящейся на стебле или корне. Разногласия касаются частных вопросов: считать ли побегами отдельные цветоножки, отдельные приросты одного и того же побега, как их называть. Мы в данной работе будем придерживаться толкования побега как стебля с сидящими на нем придатками (листьями, почками, шипами эпидермального происхождения, простыми цветоножками), образующегося благодаря деятельности одной и той же верхушечной меристемы, не подвергавшейся отмиранию.
Побег — основная, но отнюдь не самая низшая единица побеговых систем, ибо он может состоять из отдельных приростов, т. е. участков, разделенных зонами замедленного или временно прекращавшегося роста. Таковы побеги многих наших древесных пород с моноподиально нарастающими ветвями, как-то: ясеней, кленов, большинства хвойных и др. К осени на их верхушках формируются верхушечные (терминальные) почки, которые весной трогаются в рост, продолжая нарастание одного и того же побега. У многих пород верхушечные почки трогаются в рост еще в середине лета, и за один вегетационный период могут дать два или даже три прироста, как, например, у дуба. Поэтому можно назвать побег, состоящий из одного прироста (по не отдельные приросты одного побега!), элементарным или простым; побег, наросший в течение одного вегетационного периода,— годичным, побег, живущий несколько лет и состоящий из нескольких приростов,— многолетним или сложным. Естественно, разнообразие сложных побегов намного превосходит разнообразие простых, но и эти последние с трудом поддаются классификации прежде всего потому, что неизвестно, какие из их признаков считать основными, а какие — менее существенными.
Сложные побеги могут ничем не отличаться от простых, кроме своей сложности, вследствие однородности всех их приростов. Но и простые побеги могут быть не совсем просты, а также неоднородны по своей длине. Неоднородность эта может быть трех основных типов: 1) морфологическая, связанная с закономерным изменением длины междоузлий по одновершинной кривой (подробно об этом сказано у И. Г. Серебрякова, 1952, гл. VI ) и нарушением этой закономерности, когда все листья оказываются сосредоточенными у основания или вершины побега; 2) морфолого-биологическая, связанная со специализацией, степенью развития соцветий и цветков; 3) биологическая, связанная со степенью отмирания. Обычно чем более специализирован побег, тем более полно он отмирает. Однако частично или полностью могут отмирать и чисто вегетативные побеги. Особенно часто это происходит с силлептически возникшими побегами ввиду того, что их верхушечная почка оказывается не сформированной к периоду наступления холодов.
Какой-либо классификации побегов мы сейчас не имеем. Есть лишь более или менее удачные разграничения их по отдельным признакам (Нухимовский, 1970; Гатцук, 1974) или же классификации (обзоры типов) побегов у отдельных групп: родов или даже чаще всего видов растений. Не придерживаясь хронологического порядка, при котором сходные типы классификаций могут оказаться довольно далеко друг от друга, рассмотрим их в зависимости от основного подхода авторов к разграничению типов побегов: морфологического или биологического.
Морфологическая классификация. В.Н. Любименко (1900) подразделил побеги деревьев на три группы: 1) обыкновенные или длинные (с листьями срединной формации и удлиненными междоузлиями); 2) типичные укороченные (с листьями срединной формации и укороченными междоузлиями); 3) скрытые укороченные (с листьями низовой формации и укороченными меж доузлиями).
Дод ( Dode , 1905) дает следующую классификацию побегов. А. Одно-трехлетние побеги: 1) ауксибласты — ростовые побеги, появляющиеся на концах молодых ветвей; 2) мезобласты — промежуточные побеги, характеризующиеся тонкостью и более мелкими листьями; 3) хилые побеги — обычно следующего порядка. Б. Годичные побеги старых пяти-шестилетних ветвей: 4) лампробласты — массивные побеги; 5) тениобласты — тонкие побеги, возникшие летом на коротких весенних побегах; 6) брахибласты — короткие побеги старых ветвей, которые имеют вид узловатых веточек, состоящих из мелких годичных приростов. Из сложного подразделения типов побегов Дода некоторые термины прочно вошли в морфологическую литературу, например брахибласты, противопоставляемые ростовым побегам, называемым ауксибластами.
Е. Варминг ( Warming , 1908) делит побеги на длинночленистые (ассимиляционные) и короткочленистые (розеточные). Б. В. Закржевский и Е. П. Коровин (1935), И. И. Курындин и др. (1947), А. В. Гурский и др. (1953), А. А. Федоров и др. (1962) следуют в основном Доду ( Dode , 1905), деля побеги на два типа: удлиненные (ауксибласты) и укороченные (брахибласты). К брахибластам эти авторы относят укороченные цветоносные побеги многих плодовых деревьев (например яблонь» именуемых в практике садоводства плодушками или кольчатками).
Л. В. Кудряшов и Р. П. Барыкина (1970) для стланиковых кустарников выделяют четыре типа побегов: удлиненные вегетативные, полурозеточные вегетативные, укороченные (розеточные вегетативные), укороченные генеративные.
Г. Н. Зайцев (1962) выделяет следующие типы побегов жимолостей в зависимости от направления роста и расположения почек, из которых они развиваются: ортотропные годичные побеги, возникающие на адаксиальной стороне материнского побега и направленные вверх; амфитропные побеги, возникающие из трансверсальных почек и направленные примерно горизонтально; ортоамфитропные побеги, образованные из адаксиальных и трансверсальных почек; изофилъные побеги, развивающиеся обычно равномерно из всех категорий почек и расположенные под большим углом к горизонту; гипотрофные побеги, направленные вниз.
Морфолого-биологическая классификация. И. Г. Серебряков (1949) делит побеги древесных растений на вегетативные (укороченные и удлиненные — ростовые) и цветоносные, или плодущие (как правило, укороченные). Р. А. Мастинская (1971) по сочетанию двух признаков выделяет побеги ежевики Rubus caesius: 1) вегетативно-ортотропные, 2) вегегативно-плагиотропные однолетние, 3) вегетативно-плагиотронные двухлетние, 4) генеративные побеги из пазушной почки дефинитивного листа, 5) генеративные прикорневищные побеги. Биологическая классификация И. Г. Серебряков (1949) в зависимости от специализации делит побеги на три категории: слабо, неполно и крайне специализированные.
Н. Е. Булыгин (1965) классифицирует генеративные побеги на: 1) генеративные побеги, после плодоношения полностью отмирающие: а) крайне специализированные, например у вяза Ulmus scabra Mill ., б) специализированные, например у черемухи Padus racemosa (Lam.) Gilib .; 2) генеративно-ростовые, у которых отмирает генеративная часть: а) неполно специализированные, например у свидины Thelycrania аlbа (L). Pojark и клена маньчжурского Acer mandschurica Maxim, б) слабо специализированные, например у барбариса Berberis vulgaris L., в) неспециализированные, например у жимолости голубой Loniсera caеrulea L. По степени проявления вегетативной и репродуктивной функции Н. Е. Булыгин делит побеги на три основные группы: вегетативные, вегетативно-репродуктивные и строго репродуктивные.
У Е. Л. Нухимовского (1970) мы встречаем несколько подходов к разграничению побегов, например по характеру раз вития верхней части побега: 1) открытые, не заканчивающиеся соцветием, тип Loniсera caerulea ; 2) условно закрытые, заканчивающиеся кистевидным соцветием, тип Padus racemosa; 3) закрытые, у которых верхушечные меристемы образуют закрытые соцветия, заканчивающиеся терминальным цветком, тип Rosa huntica. Е. Л. Нухимовский также классифицирует побеги по степени отмирания, предлагая понятие «резид». Резид — стеблевая часть побега, которая стала многолетней после завершения им фазы видимого или верхушечного роста. В зависимости от степени отмирания резиды делятся на максимо-, медио- и миниморезиды. Побеги, не образующие резидов, т. е. однолетние, эфемерны. Побеги, образующие медио- и миниморезиды, по аналогии с эфемерными можно назвать зфемероидными.
Шпет ( Sp а t , 1912) делит побеги в зависимости от периода покоя и сформированности почек на: 1) настоящие ивановы, в которых после роста весенних побегов почка не сформирована, затем она формируется и дней через 40 начинает расти; 2) надставленные ивановы, которые возникают из терминальных конусов нарастания без образования закрытой почки и обычно без перерыва роста, редко после краткой его остановки; 3) силлептические, в которых на побеге совсем не образуется верхушечная почка и верхушка непрерывно продуцирует листья, в пазухах закладываются почки, они сразу трогаются в рост одновременно с продолжающимся ростом «обычного годичного побега"; 4) пролептические, развивающиеся на годичном побеге из уже сформированных пазушных ночек, которые находились в состоянии покоя.

Фото 1. Viburnum opulus |
Чисто биологическую, функциональную классификацию побегов ксерофитных древесных растений Каракумов дали Н.Т.Нечаева и др. (1973). В частности, у черного саксаула Haloxylon aphyllum они выделяют побеги вегетативные и генеративные; среди вегетативных побегов — ассимиляционные и ростовые, а среди ростовых — порослевые, сильные ростовые и слабые ростовые. Генеративные побеги у черного саксаула могут быть только неспециализированными, а у других древесных растений как неспециализированными, так и специализированными. Реальное разнообразие побегов гораздо более перечисленного, так как возможны самые разные комбинации признаков. Так, укороченные побеги могут быть в разной степени специализированными, отмирающими на разную величину, а удлиненные, кроме того, иметь различное направление и способ роста — прямой, вьющийся, опирающийся, цепляющийся и т. п.
Все предыдущее изложение относилось к простым или сложным, построенным из однородных приростов побегам. Но, строго говоря, абсолютно одинаковых приростов быть не может, как не может быть абсолютно одинаковых метамеров любой природы в силу закона изменчивости. Обычно приросты одного побега более или менее различаются длиной. Очень часто первые приросты бывают намного больше последующих. В других случаях неоднородность приростов одного побега может быть выражена резче. Так, укороченные побеги могут дать в последующем удлиненные приросты, что свойственно многим видам деревьев и кустарников (лиственнице, барбарису, карагане) и в той или иной мере присуще всем прочим породам с укороченными побегами. Такие сложные побеги с резко различными морфологически приростами будут в дальнейшем называться гетерогенными в отличие от побегов с более или менее одинаковыми приростами — гомогенных. Как в случае с простыми побегами, имеющими неоднородное строение, так и в случае со сложными побегами наиболее ярко гетерогенность проявляется тогда, когда побег завершается цветком или специализированным (безлистным) цветоносом. С образованием такого терминального цветоноса или цветка тесно связано последующее отмирание верхушки побега и, следовательно, остановка его роста и развития. Такие побеги И. Г. Серебряков (1952) предложил называть монокарпическими, поскольку они завершают свое развитие формированием плодов 1 и семян. |
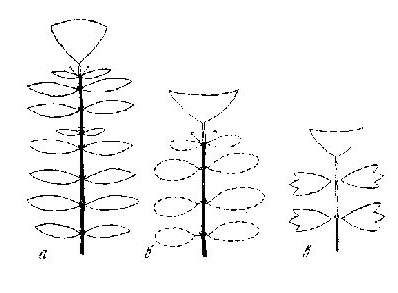
Рис. 1. Цветоносные побеги калин
а — Viburnum tinus ; б — Viburnum lantana ; в — Viburnum opulus |
Если монокарпический побег простой, состоит из одного прироста, то его называют моноциклическим, так как он имеет всего один период (или цикл) роста. Если побег сложный, то соответственно количеству приростов он может быть назван ди-, три- и полициклическим. При этом учитывается и конечный прирост, даже если он состоит из крайне специализированного соцветия. Так, например, калины Viburnum lantana и V . tinus обычно имеют дициклические побеги, но у первого вида конечный прирост его представляет собой лишь специализированное щитковидное соцветие (рис. 1, б), у второго снизу от соцветия находятся 1—2 пары листьев (рис. 1, а). И наконец, у обычной калины V . opulus соцветию так же предшествуют 2 пары ли– стьев, но побеги у нее простые, моноциклические (рис. 1, в).
В странах с умеренным и холодным климатом каждый прирост или цикл роста приходится на 1 вегетационный период. Лишь немногие породы дают по 2 или 3 прироста, да и то не совсем регулярно. Генеративные органы — специализированные соцветия — при этом несут только первые приросты. Очень своеобразна в этом отношении высокогорная среднеазиатская стелющаяся жимолость Lonicera Semenovii. Годичные побеги состоят у нее всегда из 3 приростов, т. е. трехцикличны, но лишь первый из них может нести в пазухе одной из почечных чешуй один двуцветник (рис. 2, ж). Понятие «цикличности», таким образом, применимо к любым сложным побегам, не только монокарпическим, но и чисто вегетативным. |
Побегов с «неполным циклом» попросту быть не может, поскольку один цикл — это один период роста. Неполный цикл может возникнуть лишь тогда, когда рост побега искусственно прерывается. Помимо вегетативных (акарпических) — гомогенных и генеративных (монокарпических) — гетерогенных побегов, сложными могут быть и генеративные поликарпические (Михайлова, 1972; Нухимовский, 1973). В отличие от монокарпических и подобно акарпическим они имеют неопределенное число приростов, как правило являясь полициклическими, и пазушные специализиро ванные цветоносы могут развивать ежегодно. Частным случаем поликарпического побега может считаться ди- или полициклический дикарпический побег, присущий, например, зеленчуку Galeobdolon luteum (Смирнова, Торопова, 1972).

Фото 2. Lonicera caerulea |
Если все монокарпические побеги сходны в том смысле, что имеют терминальные цветки и соцветия (различаясь типом последних), то поликарпические сильно разнятся между собой положением пазушных цветков или цветоносов, концентрирующихся у верхушек приростов, у их середины или основания. Признак этот весьма устойчив и с успехом может использоваться при характеристике таксонов разного ранга. Так, например. у циановых жимолостей подрода Periclymenum двуцветники концентрируются у концов побегов или приростов, образуя головча тое соцветие (рис. 2); у большинства жимолостей подрода — в средней части приростов (Lonicera xylosteum), у L. tatarica — ближе к верхушке, а у видов секции Caerulea распола гаются всегда в пазухах почечных чешуй в основании приростов.
Поскольку верхушка прироста дольше находится в эмбриональном состоянии, чем его основание, то чем ниже на нем располагаются цветки, тем раньше происходит их зацветание. Если побеги монокарпические, то зацветают раньше те, которые имеют более специализированные конечные приросты, а из поликарпических — с более низко расположенными генеративными побегами и цветками. Очень часто, однако, мы имеем обратную картину. Первыми зацветают поликарпические побеги с цветками, расположенными в верхней части приростов. Это происходит потому, что сформировавшиеся с осени бутоны распускаются не сейчас же, а перезимовывают. Весной следующего года они оказываются уже на приростах предыдущего, а не этого года. Подобное явление свойственно большинству наших древесных ветроопыляемых пород, а из насекомоопыляемых — волчьему лыку Daphne mezereum. Таким образом, перед нами раннее цветение фенологически, но позднее с точки зрения цикличности побегов. Субтропическая D. pontica, имеющая сходный тип приростов, цветет зимой, когда очень часто невозможно решить, что перед нами — очень позднее или, напротив, очень раннее цветение; у касандры Chamaedaphne caliculata конечные соцветия полностью сформированы уже с осени, ранней весной лишь распускаются бутоны. Здесь также моноциклические побеги с сильно растянутым периодом, протекающим между формированием цветков и их раскрыванием (Lems, 1962). |
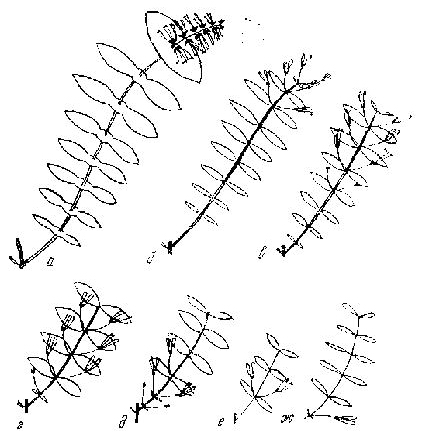
Рис. 2. Побеги ветвления некоторых жимолостей
а — Lonicera caprifolium ; б — L. japonica; в — L. tatarica; г — L. xylosteum ; д — L. caerulea; e — L. fragrantissima; ж — L . semenovii
|
Еще более интересный пример представляет собой иудино дерево Cercis siliquastrum. У него, как и у дафн, укороченные генеративные крайне специализированные соцветия образуются впервые на уже закончивших рост прошлогодних приростах зимой или ранней весной. Затем подобное цветение вследствие раскрывания все новых почек, сидящих гнездами, происходит ежегодно в течение 5—б лет. Очевидно, в этом случае можно говорить не только о поликарпичности и полицикличности побегов в целом, но и отдельных их приростов. Приросты, на которых специализированные цветоносы развиваются только один раз,— монокарпические: в первый год — моноциклические (например, большинство жимолостей), на второй — дициклические (волчье лыко), на третий, четвертый и т. д. годы — три- и полициклические; приросты, развивающие специализированные цветоносы из года в год, как Cercis siliquastrum,— поликарпические. Приросты, не развивающие генеративных органов,— акарпические. Таковы первые приросты ди- и полициклических монокарпических побегов. По аналогии с акарпическими побегами они также должны иметь свою цикличность. Заклю чаться она может, например, в длительности жизни листвы. Приросты с летнезеленой листвой — моноциклические (например, простые побеги липы, как цветоносные, так и без соцветий), с зимнезелепой — дициклические, с многолетней — полициклические.
Из приведенных примеров видно, насколько сложно явление цикличности у побегов древесных растений, ибо на их собственную цикличность может накладываться цикличность отдельных приростов, одинаковая или разная в пределах одного и того же побега. Во многом оно зависит также от того, будем ли мы рассматривать специализированные генеративные побеги в качестве самостоятельных побегов или только придатков побегов первого порядка. В первом случае понятие поликарпичности теряет свой смысл и под цикличностью приростов можно понимать только длительность жизни листвы на них. С другой стороны, в качестве придатков побегов можно рассматривать укороченные не только крайне специализированные побеги, по и менее специализированные побеги, и в этом случае явление цикличности побегов и их приростов еще более усложняется. Явление это пока нельзя считать окончательно изученным. |
Итак, признаков, по которым можно классифицировать побеги, достаточно много. Какой же из них следует положить в основу комплексной классификации побегов хотя бы древесных растений так, чтобы она охватывала все их разнообразие? Нам представляется, что принципом, который должен лечь в основу классификации побегов, должна быть та роль, которую играет тот или иной тип побега в построении кроны древесного растения, всей его сложной побеговой системы. Исходя из этого, побеги древесных (включая полудревесные) растений мы делим на следующие основные типы.
1. Первичные побеги. Они основа всей многолетней системы побегов растения, закладывающейся еще в семени. Их базальная часть всегда оканчивается корнем, и они никогда не могут быть эфемерными. Первичные эфемерные побеги свойственны только однолетним травам, да и у них свойство эфемерности относится не только к первичным побегам, но и ко всей ситстеме побегов, составляющей растение.
2. Побеги ветвления. Это боковые побеги (веточки), развниающиеся на первичном или любом другом побеге. Их роль в жизни растения весьма многообразна, но в основном сводится к увеличению вегетативной многолетней массы ассимиляционных органов (листьев или зеленых веток) и семенной продуктивности, так как именно на них находится большинство цветков и соцветий. Развиваются из обычных зимующих почек, реже — силлентически и пролептически.
3. Побеги формирования. Выполняют в основном функции омоложения побеговой системы в той или иной части или всей кроны в целом и завоевания нового прострапства. Морфологически на фоне побегов ветвления всей кроны они выделяются крупными размерами, усиленным ростом, особенно в первый, иногда и во второй год жизни, отсутствием пли сравнительно небольшим количеством цветков и соцветий. У кустарпиков именно они служат той основой, на базе которой развиваются их стволнки — главные скелетные оси. У деревьев, аэроксильных и геоксильных кустарников они вначале всегда имеют ортотропное направление роста. Они также образуют стволы у лиан, а у облигатных стлаников — их стелющиеся по поверхности земли длинные плети. Развиваются, как правило, из спящих почек, но при быстром старении побегов и их систем или при необходимости усиленного роста на первых этапах онтогенеза могут развиваться и из обычных, зимующих почек. Изредка в состояние усиленного роста переходят укороченные побеги, как, например, у многих хвойных, особенно у лиственницы, и некоторых лиственных: берез, тополей; п др. Соответственно со своими основными функциями или морфологическими особенностями разными авторами назывались по-разному (табл. 1). Далее мы часто будем пользоваться термином «турион».
4. Побеги дополнения. Морфологически во всем походят на побеги ветвления, но роль их сходна с ролью побегов формирования. Они могут появляться из дополнительных и спящих почек в качестве реакции на старение или повреждение растения, но свойством усиленного роста не обладают. Изредка имеют весьма специальное назначение, развиваясь и виде колючек или укороченных цветоносных побегов (каулифлория).
Развитие из спящих почек побегов как формирования (турионов), так и дополнения стимулируется не только собственно старением, но и такими чисто механическими факторами, как обрезка, обламывание и изменение положения ствола или ветви с вертикального на наклонное и тем более горизонтальное. При этом просыпаются и трогаются в рост только почки, расположенные на стороне, обращенной вверх, и развивающиеся из них побеги, таким образом, как бы стремятся восстановить «вертикальность» наклонившейся оси.
5. Скрытые побеги — долго живущие спящие почки и продукты их ветвления. Подрастая ежегодно на величину годичного прироста древесины спящие почки превращаются в настоящие, хотя и укороченные, побеги (Любимепко, 1900). Обычно скрыты в коре, образуя более или менее крупные скопления
(сферобласты), и играют роль резерва, на основе которого могут развиваться побеги двух предыдущих типов, но в «полускрытом» виде могут выступать и самостоятельно, выпуская как бы прямо из ствола несколько листиков (каулифилия) или неолиственных цветоносов (каулифлория).
6. Эфемерные побеги — однолетние. По своему виду походят на побеги как ветпленпя, так и формирования, развиваются из почек всех типов, в том числе, и притом наиболее часто, из не зимующих, силлептически и пролептически. По некоторым своим свойствам могут быть распределены на группы, соответствующие второй, третьей и четвертой группам данной классификации, но всех их объединяет одна важная черта: они не играют никакой роли в построении многолетней системы побегов древесного растения (не участвуют в построении его «базы»), что для этого последнего, разумеется, далеко немаловажно. В основном играют роль обогащения кроны ассимиляционным и репродуктивным аппаратом.
Эфемерность побегов, как и эфемероидность, может быть факультативной и облигатной. Факультативная эфемерность возникает из-за полного отмирания тех или иных обычно многолетних побегов в особо суровых условиях существования в особо суровые зимы, или вследствие общего ослабления растения в конце онтогенеза, или вследствие нападения вредителей и т. п.
Между всеми перечисленными тинами побегов имеются, разумеется, как и в любой другой классификации, переходы. Особенно часты они между побегами формирования и дополнения, а также между побегами ветвления и эфемерными побегами. Положение осложняется тем, что некоторые типы побегов могут переходить в другие в процессе их собственного развития. Так, свойство усиленного роста могут приобретать все типы побегов (кроме эфемерных), побеги же формирования в конце своего развития могут замедлять свой рост. Какому принципу классификации надо следовать в этом случае? По нашему мнению, первичный побег, как бы он ни изменял темп своего роста, всегда остается первичным побегом, основой дебетовой системы всего растения. В других же случаях следует отдавать предпочтение свойству усиленного роста. Так, побеги ветвления, дополнения и скрытые превращаются иногда в побеги формирования, скрытые побеги — в побеги дополнения, укороченные побеги ветвления — в длинночленистые побеги ветвления. Ниже рассмотрим каждый из выделенных типов более подробно.
При дальнейших подразделениях названных выше типов побегов мы следуем тому же принципу важности роли, которую играет та или иная особенность побега в построении кроны древесного растения. Важность эта возрастает пропорционально размерам побегов, что при прочих равных условиях зависит от: 1) числа их приростов, т. е. степени сложности; 2) степени «розеточности» приростов; 3) степени их эфемероидности (обратно пропорциональная зависимость). Число приростов, в свою очередь, зависит от степени цикличности; чем цикличность выше, тем больше число приростов в побеге, а так как акарпические и поликарпические побеги обладают большей в среднем цикличностью, чем монокарпические, то за открытыми побегами надо признать более важную роль в построении побеговых систем. В целом наиболее целесообразной представляется нам следующая иерархия признаков, которая может служить основой для классификавсех выделенных типов.
I. Побеги сложные: 1) удлиненные и полурозеточные: а) откытые (вегетативные и генеративные), б) закрытые (те и другие далее делятся на группы соответственно своей цикличности); 9) розеточные: а) открытые, б) закрытые.
II. Побеги простые: А) неэфемероидные (образующие максиморезиды): 1) удлиненные и полурозеточные: а) открытые (вегетативные и генеративные), б) закрытые; 2) розеточные: а) открытые, б) закрытые; Б) эфемероидпые (образующие медио- и минимо-резиды) — по-видимому, только удлиненные и полурозеточные: а) открытые, б) закрытые.
Первичные побеги, а также побеги ветвления и дополнения имеют все градации данной шкалы. Побеги формирования в морфологическом плане можно было бы именовать сверхудлиненными. Они не имеют градаций, соответствующих розеточному типу. Их место занимают побеги с корневищным, подземным основанием, свойственные геоксильным кустарникам, очень короткие у одних видов (например, лещина Corylus avellana L .) и длинные у других (шиповник Rosa ciimamomea L.). Называются они ксилоподиями ( Du- Rietz , 1931) или ксилоризомами (Дервиш-Соколова, 1966).
Кроме приведенной классификации побегов по их биоморфологическим признакам, возможны и классификации по другим признакам, как-то: 1) производящие побеги почки: а) биологические свойства (таковы выше приведенные градации Шпета), б) положение (придаточные, пазушные, корневые и т. д.); 2) взаимное положение побегов (см. следующий раздел); 3) направление и способ роста — ортотропные, ползучие, простратные, обвивающие и.т. д. (см также классификацию Зайцева); 4) возраст — многолетние, однолетние (первого года развития), годичные, весенние, осенние; 5) находящиеся на побегах придатки - колючие, гладкие, шероховатые, с супротивным или очередным листорасположением и т. д.
Основные системы побегов
Системы побегов, образующих крону древесного растения (кроны, образованные всего одним побегом, таи называемые пальмовидные, присущи только древовидным папоротникам, саговникам, пальмам и еще небольшому числу древесных однодольных, оби тающих в тропиках), могут возникнуть не иначе, как только путем их ветвления. Наиболее распространенные типы ветвления — моноподиальное, т. е. одновременное с ростом основного, материнского побега первого порядка, и симподиальное, следующее только после завершения его роста. Наиболее сильные (длинные и долговечные) боковые дочерние ветви (или конечного его прироста), либо в средней или нижней его части. В первом случае говорят об акротонии, во втором — о мезотонии и в третьем - о базитонии боковых побегов.
Частным случаем акротонного снмподиального (акросимподиального, по Л. Е. Гатцук, 1970) ветвления является ложнодихотомическое ветвление, при котором рост материнского побега (первого порядка) продолжают 2 побега, развивающихся у самой его верхушки, так что она часто бывает почти незаметна. Иногда же рост материнского побега продолжает всего 1 дочерний побег, и тогда совокупность 2 и более побегов бывает очень сходна с совокупностью приростов 1 побега или даже более того с совокупностью узлов и междоузлий 1 прироста. Подобный случай представляет собой стебель многих виноградов, настоящий побег которых всегда заканчивается усиком, а побег, продолжающий его рост, развивается из пазухи единственного листа. Здесь уместно, очевидно, говорить о ложных побегах, являющихся на самом деле совокупностями простых побегов все более высокого порядка, но биологически все же вполне аналогичных настоящим побегам. Побег, продолжающий нарастание материнского побега, носит название побега продолжения (Troll, 1937). Наиболее сильные побеги, возникающие при симподиальном ветвлении или при моноподиалыюм в том случае, если дочерние побеги (один или несколько) перерастают материнский,— побеги замещения.
Такие термины, как побеги продолжения, замещения, материнские, дочерние первого, второго и более высоких порядков, лидерные (наиболее сильные из замещающих), боковые, верхние или нижние и некоторые другие, характеризуют не сами побеги, а их взаимное положение, возникающее в результате ветвления. Все эти их свойства, естественно, не могут быть использованы в приведенной выше классификации побегов по морфо-биологическим признакам.
Совокупность материнского и дочерних побегов уже представляет собой простейшую систему. Частный ее случай — «система скелетного моноподиального побега — скелетный моноподиальный побег вместе со всеми возникшими на нем цветоносами и моноподиальными побегами обрастания и обогащения» (Гатцук, 1974а, с. 107), т. е., согласно этому автору, простейшую систему побегов образует долговечный побег с сидящими на нем эфемерными побегами — генеративными и вегетативными. Метамерная единица следующего порядка — это совокупность побегов, образующих всю крону дерева или стволика куста,— «система моноподиальных побегов главной оси» (Гатцук, 1970, с. 108).
Однако между одним долговечным побегом и их множеством, образующим крону, дистанция столь огромного размера, что невольно возникает мысль о существовании иных систем побегов, более высокого ранга, чем «система скелетного моноподиального побега», и более низкого, чем вся крона. Системы, образуемые различного рода побегами, как и сами эти побеги, весьма различны и, следовательно, играют разную роль в построении кроны древесного растения, одни — более, другие — менее важную. Так, очевидно, что основу «скелета» кроны составляют неспециализированные удлиненные побеги, а не укороченные (если, конечно, они не единственный вид побегов, образуемых растением). Скрытые побеги и системы скрытых побегов вовсе не принимают участия в построении кроны, являясь лишь резервом, пополняющим ряды побегов дополнения и формирования. Эфемерные побеги и их системы (возникающие силлептически и пролептически) — лишь временные поселенцы в кроне, не участвующие в создании ее многолетней части, «скелета».
Напротив, без первичных побегов и их систем не может быть и самого растения. Они фундамент для других побегов и систем побегов. У одних растений первичные побеги или замещающие их системы существуют очень долго, зато у других они крайне недолговечны. У многих геоксильных кустарников они живут всего около 10 лет, что при общей продолжительности жизни этих кустарников более 100 лет, конечно, срок небольшой. Однако где проходит граница между системой первичного побега и последующими системами и, вообще, что надо считать за основную единицу систем побегов?
Как мы видели выше, среди всех типов побегов побеги формиронания занимают особое место. Во-первых, именно благодаря им образуется сама кустовидная форма роста, почему они и носят часто название побегов кущения или возобновления. Во-вторых, именно они формируют в основном крону куста благодаря своим крупным размерам (сверхудлипенности), из-за чего часто называются ростовыми побегами или побегами «нарастания». И наконец, в третьих, на них в основном лежит миссия обновления стареющих систем побегов как отдельных частей кроны, так и в целом всей кроны, о чем свидетельствуют такие их наименования, как побеги «возобновления», «обновления», «замещения» (табл. 1). Очевидно, побег (и система, им образуемая), приходящий на смену отмирающей системе, не может принадлежать ей же, а играет уже самостоятельную роль, имея, так сказать, совершенно противоположную, «омолодительную», а не «стареющую» физиологию, что морфологически и биологически выражается в явлении усиленного роста. Поэтому не будет, повидимому, большой ошибкой считать, что «правом» образовывать свои автономные системы обладают три типа побегов: первичные побеги, побеги формирования и дополнения. У кустарников, особенно геоксилытых, кустящихся на уровпе почвы и ниже, совершенно выдающаяся роль при этом принадлежит побегам формирования. Собственно, кустарниковая форма роста представляет собой совокупность систем побегов формирования (СПФ) в пространстве и смену этих систем во времени.
Под «автономными» мы понимаем такие системы, которые не являются частями других систем того же уровня полимеризации метамеров. Если побег — это первый уровень, основной блок метамерной системы тела всех семенных растений, то система первичного побега (СПП) и СПФ — второй.
Степень автономности может быть, естественно, различной. Так, несомненно, СПФ, развивающиеся в основании куста, а тем более с ксилоризомами, обладающими своими корневыми системами, более автономны, чем СПФ в корне. Системы побегов дополнения (СПД) менее автономны, чем СПФ, развивающиеся в корне, о чем свидетельствуют и их «нормальные» или даже укороченные размеры. Поэтому СПД, как и системы побегов ветвления (СПБ), развивающиеся в кроне, мы считаем не равноценными СПФ, а лишь частями их или СПП. Самостоятельное значение они приобретают тогда, когда ветвь (СПП) укоренится и сможет существовать вполне автономно. Тогда СПВ и СПД выходят на тот же уровень полимеризации, что и СПП и СПФ.
К третьему уровню полимеризации побегов принадлежат системы, образованные самими СПФ, т. е. «системы систем» — слагающие куст стволики (основные или главные скелетные оси, по П. Г. Серебрякову, 1962), к четвертому — кусты в целом и, наконец, к пятому — совокупности таких (первичного и парциальных) кустов, создающие клон.
Автономность СПФ, возникающих даже в кроне деревьев, хорошо подтверждается тем, что на фоне всех ветвей кроны, являющейся СПП, они выглядят как инородное тело, развивая свою собственную крону, часто в виде миниатюрного деревца, подобного материнскому, со своим стволом, растущим ортотропно и образованным побегом формирования (настоящим или ложным) и горизонтально распростертыми ветвями, образованными СПВ. Размер таких «деревцев» сильно варьирует от очень крупных, достигающих нескольких метров (развиваются они, как правило, на горизонтальных сучьях и наклоненных стволах, рис. 3), до довольно мелких, всего в 10-20 см, развивающихся, как правило, на мелких сучьях и вертикально стоящих стволах (рис. 4).

Рис. 3. Две системы формирования в кроне тунга
|
Кроме систем, образующих соподчиненную иерархию, можно выделять комплексы побегов. В системе все побеги тесно связаны между собой, и, естественно, без побега формирования или главной скелетной оси не может быть СПФ или СПП. Комплексы же продолжают существовать при удалении любого побега, образующего его (если их, естественно, больше двух). Такие комплексы образуются, например, при развитии в побеги всех пазушных сериальных или коллатеральных почек, или при ветвлении спящих почек, или при любом другом скученном расположении побегов.
В некоторых случаях побеги в комплексе образуют весьма естественные группы, например тогда, когда они развиваются из сериальных или коллатеральных почек, находящихся на одном узле. Очень часто в таком комплексе находятся цветоносы и вегетативные побеги.
Итак, основной единицей побегового тела кустарника является система побега формирования (СПФ), поэтому основой морфогенеза кустарников должно быть развитие этой структуры во времени — основной цикл (Лучник, I960).
Система побегов формирования разных видов и особенно жизненных форм отличаются друг от друга размерами побегов формирования и ветвления, направлением их роста, степенью специализации, расположением побегов ветвления на побеге формирования, количестпом порядков побегов ветвления, наличием и особенностями побегов дополнения. |

Рис. 4. Система побега формирования на стволе платана |
У разных видов основные циклы как понятие временное различаются главным образом длительностью своих периодов. Так как в начале преобладают процессы роста, затем ассимиляции и плодоношения и в конце отмирания, мы выделили три периода: I — период преимущественного роста («рост»); II — преимущественная ассимиляция и плодоношение («стабилизация», пли «кульминация»), который в зависимости от длительности основного цикла может быть растянутым или кратким, совпадая с его пиком; III — «отмирание», в течение которого, наряду с продолжающимися процессами роста, ассимиляции и плодоношения, происходит все большая интенсификация процессов отмирания побегов, что ведет к снижению интенсивности всех прочих функций и в конце концов к полному или почти полному отмиранию всей СПФ, хотя не менее половины побега формирования остается иногда живой и входит в систему побегов более высокого ранга.
В большинстве случаев переход одного периода в другой происходит плавно и развитие некоторых порядков побегов ветвления совпадает с периодами «рост» и «стабилизация», а на нисходящей стадии — с периодами «стабилизация» и «отмирание». Поэтому основной цикл и его периоды можно измерять как в единицах временных (год, месяц, вегетационный период), так и. порядками побегов ветвления. Рассмотрим периоды основного цикла более подробно. I период — «рост» - начинается с момента образования побега формирования из спящей почки, В типичном случае, например у чубушников, жимолостей, большинства шипопников и др., он длится один вегетационный период, в течение которого побег формирования достигает значительной величины. У некоторых других кустарников этот период более долгий: например, у многих рододендронов, Viburnum awabuci, V. tinus, V. japonicum, Hibes triste период «рост» охватывает время разлитии побегов формирования и ветвления первого — четвертого и более порядков, если они отходят от нижних приростов побега формирования. II период — «стабилизация» или «кульминация» - начинается обычно с роста побегов ветвления первого - третьего порядков. У кизила, рододендронов, жимолостных — плейокормных деревьев и аэроксильных кустарников он длительный и может продолжаться во время развития побегов ветвления второго - десятого порядков. У типичных геоксильных кустарников этот период более короток и приходится на развитие побегов ветвления второго - четвертого порядков. У многих видов именно в этот период активно образуются побеги дополнения. III период - «отмирание» - начинается у чубушников, форзиции, бузин, жимолостей и многих других уже при развитии побегов ветвления третьего порядка; у древесных жимолостных он не резко отграничен от предыдущего. У гордовины в основном цикле до 15 порядков побегов ветвления, однако уже с четвертого - шестого порядка количество побегов ветвления не увеличивается. |
В дальнейшем начинается отмирание побегов конечных годичных приростов и, когда образуются побеги ветвления седьмого-восьмого порядка, количество вновь образовавшихся и отмерших побегов становится разным. Затем число отмерших побегов все быстрее увеличивается по сравнению с образовавшимися, так как начинают отмирать не только конечные приросты, но и предыдущие. В период «отмирания» на ослабленных ветвях нерегулярно появляются побеги дополнения, подобные ослабленным побегам ветвления, всегда без цветков или соцветий. За счет их развития основной цикл продлевается иногда до 10 лет и более.
У бузины Sambucus racemosa верхняя часть побега формирования отмирает уже к концу первой вегетации. На второй год отмирает значительная часть побега формирования, побеги ветвления также отмирают на две трети. К концу основного цикла функционирующей остается только половина побега формирования, до места образования на нем побега формирования следующего порядка.
Таким образом, СПФ отмирают не полностью. Нижняя часть побега формирования становится многолетней; на ней появляется дочерний побег формирования, нижняя часть которого со временем также становится многолетней.
Чем выше в кроне, тем больше образуется дочерних систем побегов формирования, но более мелких, со все быстрее проходящим основным циклом и все более сильным отмиранием. Тут проявляется как бы равновесие между отмиранием и появлением новых систем побегов. Жизнедеятельность куста в целом и его метамерных осей представляет собой как бы борьбу между процессом обновления и отмирания.
Периоды основного цикла в какой-то мере можно сравнить с виргинильным, генеративным и сенильным периодами большого жизненного цикла онтогенеза. Вначале в онтогенезе преобладают процессы роста (виргинильпый период), затем в генеративном периоде плодоношения и в сенилыюм периоде отмирания. По И. Г. Серебрякову (1952), водяные побеги (побеги формирования) обладают рядом свойств молодых растений.
Периоды основного цикла изменяются в зависимости от возрастного состояния и экологии вида. В начале онтогенеза, когда все ростовые процессы усилены и основной цикл сокращен, преобладает период «рост», процессы отмирания, хотя и имеются, но выражены слабо. В средневозрастном генеративном состоянии периоды основного цикла выявляют свои ритмы с наибольшей полнотой. В сенильном периоде система побегов стареет, период «отмирание» может быть растянутым и длиться несколько лет. Периоды основного цикла изменяются и в зависимости от местообитаний, особенно в суровых природных условиях.
От периодов основного цикла зависит строение СПФ, в частности расположение и цикличность побегов ветвления. В начале основного цикла на побеге формирования побеги ветвления первого порядка развиваются акросимподиально в его верхней части, а почки в средней и нижней частях побега формирования остаются спящими. Побеги ветвления, как правило, располагаются в его верхних 4-5 узлах, например у жимолости кавказской (Lonicera cancasica), чубушника (Philadelpdus coronarius), азалии поптийской (Azalea ponlica), рододендрона мелколистного (Rhododendron parvifolium), физокарпуса (Physocarpns opulifolia), или только в 1, реже в 2 верхних узлах побега формирования, например у свидипы (Thelycrania alba), калины лавролистной (Viburnum tinus), сирени (Syririga vulgaris). Сильнее эта особенность выражена у жимолости Карелина (Lonicera karelinii), у которой побеги ветвления первого порядка образуются из 2 боковых почек, формирующихся рядом с терминальной. Реже встречаются виды, побеги ветвления которых располагаются по всей длине побега формирования. К такому типу относится жимолость блестящая (Lonicera nitida), а так же, например, кизильник (Cotoneastcr melanocarpa), спирея средняя (Spirea media), у которых побеги ветвления образуются акросимподиально, но также встречаются в средней и нижней частях побега формирования. Для видов, у которых побег формирования ко времени образования побегов ветвления стал плагиотропным, побеги ветвления располагаются на его верхней стороне, тогда как на нижней стороне они ослаблены или их нет вовсе. Это особенно характерно для вьющихся жимолостей, например для жимолости японской и каприфоли, а также для стелющихся, например для жимолости Семенова, реже для прямостоячих (Зайцев, 1962).
У некоторых видов намечается закономерность в размещении вегетативных и генеративных побегов ветвления. Например, у Spirea media, калины обыкновенной (Viburnum opnlus), калипы восточной (V . orientale) вегетативные побеги ветвления с 3-5 парами листьев располагаются в средней части побега формирования, а генеративные — в верхней его части с 2-3 парами ассимилирующих листьев ниже соцветия. У бузины красной вегетативные побеги ветвления располагаются в средней части побегов формирования с 3-6 парами ассимилирующих листьев, генеративные — в верхней части побега формирования с 2 парами ассимилирующих листьев.
У видов с побегами ветвления, дифференцированными па вегетативные и генеративные, первые всегда длиннее вторых и имеют большее количество пар листьев. У кустарников с открытыми лобегами (крыжовников, смородин, жимолостей и др.) нет резкой разницы между вегетативными и генеративными побегами ветвления, так как число соцветий может сильно колебаться. Обычно побеги без соцветий — ослабленные, затененные веточки, а побеги с наибольшим количеством соцветий находятся на наиболее освещенных местах кроны.
У некоторых видов резко различаются побеги ветвления разных порядков. Например, у жимолости кавказской побеги ветвления первого порядка в основном без двуцветников, в чем они сходны с побегами формирования. С увеличением порядка размер и количество узлов на них уменьшаются. Приводим примеры типичных побегов ветвления первого порядка некоторых жпмолостей (рис. 2).
Величина и количество междоузлий побегов ветвления более постоянны, чем те же параметры побегов формирования.
У видов с полициклическими побегами с течением основного цикла изменяется и цикл побегов ветвления либо в сторону уменьшения, либо увеличения. Например, у калины Авабука (V. awabncki) сначала побеги полицикличеокие, затем начинают преобладать ди- и трициклические, а в конце и моноциклические. У калины лавролистной (V . tinus) в начале цикла побеги ветвления ди- и трициклические, а в конце — ди- и моноциклические. У рододендронов и других вересковых с 3-5 циклическими побегами чем ближе к концу основного цикла, тем более возрастает цикличность отдельных побегов (до 10 лет и более).
Как правило, побеги ветвления выполняют функции плодоношения, фотосинтеза и ассимиляции. Но у некоторых видов побеги первых порядков морфологически переходны к побегам формирования, выполняя, очевидно, также и ростовую функцию, как, например, у жимолости кавказской, дёрена белого, саксаулов белого и черного. У жимолости Маака побеги ветвления первого порядка также без двуцветников, достигают 40-50 см, имея 8-10 узлов. У Philadelpnus coronarius (Москва, Останкино) побеги ветвления первого порядка — 50-70 см, второго — 3О-40 см, третьего — 15-20 см; у гортензии раскидистой (Hydrangea paniculata) (Москва, ГБС) побеги ветвления первого порядка — 50-60 см, второго — 35-40 см. Побеги ветвления последних по рядков СПФ у многих кустарников также не несут цветков и соцветий и не выполняют, таким образом, функции плодоношения.
Каждый вид в определенном местообитании и возрасте имеет определенное количество порядков побегов ветвления, что являетcя характерной его особенностью. Например, у калины лавролистной их 10—16, у жимолости Карелина — 5-7, жимолости Глена — 4-5, жимолости душистейшей — 4, жимолости голубой — 3-4, бузины кистистой, жимолости японской и снежноягодника белого — 2-3, бузины канадской— 1-2, бузины травянистой — 1, у Hydrangea opuloides (Батумский ботанический сад) и у Thelycrania alba (Москва, ГБС) в системе побега формирования 4-5 порядков; у дейции шероховатой (Deutzia scarba) (Батумский ботанический сад) — 3-4 порядка; у шиповника иглистого (Rosa acicularis) (с. Мопды Бурятской АССР) — 3 порядка и т. д.
У большинства кустарников жимолостных количество порядков колеблется в пределах 3-4, у плейокормных деревьев мо жет достигать 20, а у трав — всего 1 . В крайних условиях высокогорий количество порядков может быть большим. Так, для жимолости Турчанинова среднее число порядков побегов ветвления в таежном поясе — 3-4, а в высокогорьях Восточного Саяна — 20 и более.
Периоды основного цикла влияют на свойства побегов формирования, главным образом на продолжительность и направление их роста. Вначале побег формирования ортотропный, но затем направление его роста и побегов ветвления постепенно изменяется. В период стабилизации верхняя часть системы побега формирования дуговидно изгибается. Особенно наглядно это видно на примере крыжовников, чубушников, физокарпусов, жимолостей кавказской, монетолистной, Глена и многих других.
В дальнейшем под тяжестью побегов ветвления побег формирования постепенно приобретает плагиотропное направление роста, ортотропной остается лишь его нижняя часть. У шпалерных кустарников побеги формирования более длинные, стелющиеся, ортотроппо растущие только в самом начале своего развития. У лиановых жимолостей побег формирования может менять направление своего роста неоднократно, что зависит от наличия или отсутствия опоры.
Очень существенным признаком СПФ, определяющим многие ее свойства, является длительность роста побега формирования, так как именно он — основа всей системы. В зависимости от длительности нарастания побега формирования СПФ можно подразделить на следующие группы (рис. 5):
а) моноподиальную, когда нарастание побега формирования потенциально продолжается весь основной цикл; как, например, у линией, кассиоп, караганы гривастой, смородин;
б) моноподиалъно-симподиальную, когда нарастание побега формирования иногда продолжается долго, но не весь основной цикл, а может и прекратиться на второй-третий год; как, например, у многих шиповников, рододендронов, багульников, жимолостей татарской и Глена;
в) симподиальную, когда побег формирования состоит всегда из одного элементарного побега, на второй год основного цикла нарастания его никогда нет (бузина кистистая, снежноягодник, леспедеза двуцветная, чубушник, сорбария и др.).
|

Рис. 5. Три основные группы СПФ.
а — моноподиальная: б — моноподиально-симподиальная; в — симподиальная |
Две последние группы разделяются на две подгруппы каждая: а) вегетативную, с открытыми побегами формирования — терминальная почка на них всегда остается вегетативной (чубушник, кассиопа, жимолости, бузина кистистая), б) генеративную — нарастание побега формирования прекращается вследствие заложения терминального соцветия (большинство калин, снежноягодники, лейцестерия, рододендроны, багульники, шиповники и др.).
У различных видов основной цикл протекает в разные сроки. В северных районах умеренного пояса этот срок ограничен сезонностью климата. Каждую вегетацию развивается 1 порядок побегов, хотя иногда в благоприятные годы возможно образование пролептических или Ивановых побегов.
У некоторых видов уже облигатно за один сезон разливается 2-3 прироста. Например, у высокогорной жимолости Семенова (в культуре, ГБС) побеги формирования и ветвления дают 2-4 прироста за вегетацию, а основной цикл длится 3 года. У дёрена белого, леспедезы двуцветной, интродуцированной в ГБС среднеазиатской жимолости монетолистной побеги ветвления первого порядка развиваются большей частью в год роста побега формирования. На следующий год появляются побеги ветвления второго и третьего порядков. Система побега формирования у жимолости монетолистноп состоит из 3 порядков побегов ветвления, а основной цикл длится 2 вегетационных периода. Еще более ускорен основной цикл у субтропической жимолости японской, которую мы наблюдали в Батуми, где она дичает. СПФ ее имеет 3, реже 4 порядка побегов ветвления, а весь основной цикл проходит в течение 1 вегетационного периода. Ускоренные ритмы основного цикла характерны для южных видов, например гортензии Hydrangea opuloides (Батумский ботанический сад), мирикарии Myricaria alopecuroides (Таджикская ССР, окр. Гарма). Но у субтропической жимолости душистейшей и жимолости Стендиша |
ежегодно появляется только 1 порядок побегов ветвления, а цветут они зимой, в декабре-январе. В зависимости от длительности мы предлагаем выделить у кустарников следующие три группы основных циклов: длительностью 10 и более лет; длительностью 3-10 лет; длительностью 1-3 года.
Первый тип характерен только для деревец, аэроксильных и крупных геоксильных кустарников, у которых турионы морфологически походят на побеги ветвления первых порядков; величина побегов в продолжении основного цикла постепенно падает. Второй тип СПФ характерен для аэроксильных и геоксильных кустарников с регулярными сменами. Системы побега формирования располагаются в кроне и в основании главных скелетных осей. Турионы специализированы и четко отличны от побегов ветвления разных порядков, хотя у некоторых типов имеются плавные переходы от побега формирования к побегам ветвления. У большинства видов побеги моноциклические. Третий тип СПФ характерен для низкорослых геоксильных кустарников, развивающих, как правило, подземные побеги (снежноягодники, бузины, сорбарии). Новые СПФ образуются в основании недолговечных скелетных осей или от подземных ксилоподиев (ксилоризом).
Cкелетные оси
Термины «скелетные ветви» и «скелетные сучья» были использованы впервые плодоводами (Шитт, 1952) для характеристики наиболее долговечных частей кроны плодовых деревьев. Такой же неопределенный характер он носит и у морфологов, что справедливо отметила Гатцук (1970). Мы под скелетными осями будем понимать все многолетние ветви, а под главными скелетными осями — стволы, стволики и крупные сучья деревьев. Характерная черта кустарников та, что они в отличие от де ревьев имеют не один крупный ствол, а несколько более мелких стволов или стволиков — главных скелетных осей, связанных основанием друг с другом и образующих как бы пучок - куст. Величина растения при этом не играет существенной роли. Кусты могут быть «древовидными», мало в чем уступая деревьям по размерам, как, например, образуемые березами и другими древесными породами при срубании ствола и росте новых стволов из водяных побегов. В этом случае говорят обычно о многоствольных деревьях, но это все же кустарники, образующиеся только не с самого начала онтогенеза растения, а лишь с его середины или уже к его концу. Вторичные стволы весьма похожи на первичный, но все же меньше по размерам и менее долговечны. По существу, развиваясь из водяных побегов, т. е. из побегов формирования, они представляют собой СПФ, только с достаточно длительным, в несколько десятков, а может быть и до более чем 100 лет, циклом развития. И хотя для большинства древесных пород умеренной полосы характерно явление смены крон, т. е. замены отмирающих сучьев новыми, развивающимися из СПФ, тем не менее их ствол образован обычно системой первичного побега или одной СПФ. У типичных же геоксильных кустарников их стволики (главные, или основные скелетные оси) всегда построены из нескольких СПФ. Как происходит их формирование, легко проследить на примере широко распространенных в культуре спирей, чубушников, жимолостей, калин.
Многолетние стволики (главные скелетные оси) геоксильного кустарника отходят от его основания и состоят как бы из отдельных этажей — ярусов. В нижней части стволики или ветви каждого яруса имеют ортотропное направление, а в средней — дуговидное или несколько плагиотропное. Это объясняется тем, что побеги формирования, из частей которых возникла главная скелетная ось, сначала растут вертикально, а затем их верхушки дуговидно изгибаются. На плагиотропной части побега формирования размещаются дочерние побеги формирования, более молодые и потому менее толстые по сравнению с материнским, а еще выше на плагиотропной части дочерней оси — оси следующего порядка, более тонкие и меньшие по размеру, чем предыдущие и т. д. Поэтому многолетние скелетные оси создают этажный силуэт кустарника, все уменьшаясь снизу вверх, по становясь все многочисленнее (рис. 6).
|
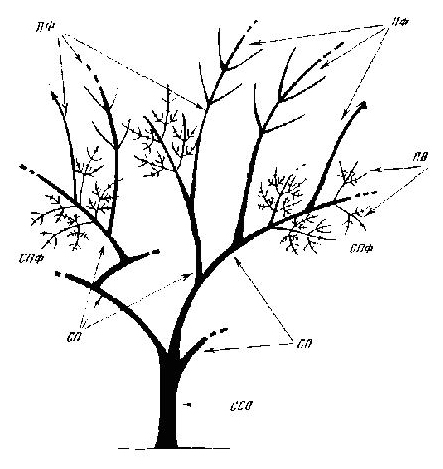
Рас. 6. Строение системы составной скелетной оси
ПФ — побеги формирования; ПВ — побеги ветвления; СПФ — системы побегов формирования; СО — скелетные оси; ССО — составные скелетные оси |
Процесс образования главной скелетной оси начинается с роста от основания куста мощного побега формирования, часто достигающего полной высоты кустарника. На второй - третий год вырастают побеги ветвления, а верхняя часть побега формирования обычно наклоняется горизонтально. Иногда уже на второй год, но чаще на третий - четвертый, от средней или нижней его части отходит побег формирования следующего порядка. В это время в верхней части материнской СПФ побеги ветвления начинают постепенно отмирать, и вскоре вся верхняя часть СПФ отмирает до места отхождения первого (сверху) дочернего побега формирования. После этого разрушение материнской системы приостанавливается, начинается отмирание системы первого дочернего побега формирования.
Дочерние побеги формирования, развивающиеся в верхней части материнского побега, мелкие, с быстропроходящим циклом. После отмирания очередной дочерней системы гибнет часть материнского побега формирования до места отхождения последующей (сверху вниз) дочерней СПФ, отмирание которой длительнее, чем предыдущей, поскольку она более мощно развита. В то время, когда гибнет система материнского побега формирования, дочерние системы, расположенные ниже, могут уже образовывать собственные дочерние системы (третьего порядка). Таким образом, волна отмирания как бы сталкивается с волной роста и нижняя часть материнского побега формирования остается живой до места отхождения одной из наиболее сильных дочерних СПФ, которая отходит от материнской в месте ее перегиба. Утрачивается горизонтальная часть СПФ, а вертикальная, представленная, как правило, первым годичным приростом или его частью, остается живой довольно долго, входя в состав метамерной системы более высокого порядка. |
Из всего этого можно заключить, что так называемые скелетные оси кустарников (Серебряков, 1955, 1962), т. е. их стволики, отличаются от скелетных осей (стволов) деревьев не только меньшей продолжительностью жизни, но и более высокой степенью полимеризации побеговых систем. Если у деревьев их стволы образованы системой первичного побега, то стволики кустарников построены частями СПФ, т. е. они не могут быть гомологами первых. Стволики кустарников — ложные стволики, похожие на стволы деревьев лишь внешне. Следовательно, они заслуживают и другого названия — сложных или составных главных скелетных осей (ССО). Составные скелетные оси, как сказано отчасти выше,— совокупности нижних частей побегов формирования все более возрастающего порядка. Система ССО — все СПФ, участвующие в их построении, а также и все другие побеги — формирования, дополнения и ветвления, включая и эфемерные, располагающиеся на стволиках и их ветвях.
Количество порядков СПФ в составе сложной скелетной оси, высота и долговечность ССО зависят от местообитаний (Серебряков, 1962; Шафранова, 1964, 1968; и др.). В одних местообитаниях мы находим длительно живущие системы скелетных oceй, в других они отмирают быстрее. Побеги формирования и ветвления, СПФ, скелетные оси разных порядков, ССО (стволики) имеют разное количественное соотношение у разных кустарников. У большинства видов побегов ветвления сотни, побегов формирования несколько десятков, дочерних скелетных осей десятки и главных скелетных осей (стволиков) всего несколько, единицы.
Каждому виду присущи свои особенности ССО. Во многом это зависит от длительности основного цикла, степени отмирания СПФ. Например, в основном цикле у бузины красной отмирание сильно выражено уже на второй - третий годы. К моменту развития дочерних СПФ вторых - третьих порядков верхняя часть системы материнского побега формирования полностью отмирает, а многолетняя вертикальная часть (его основание, стволик) остается оголенной. Происходит интенсивное очищение кроны, поэтому бузина, особенно в конце онтогенеза, выглядит часто одноствольным деревцем, построенным из не очень резко выраженных ярусов. Отмирание систем побегов у других видов, например у жимолости съедобной, выражено слабее, и в кроне кустарника всегда скапливаются отмершие или почти отмершие верхушки мелких СПФ разных порядков.
|
Таблица 2
Характеристика главных скелетных осей некоторых кцстарников в условиях интродцкции (Москва, Главный ботанический сад)
Вид |
Основной цикл , годы |
Высота, м |
Количест– во ярусов скелетных осей |
Количество главных скелетных осей |
Syringa vulgaris |
7-10 |
2,0-2,5 |
5—7 |
5-10 |
Thelycrania alba |
7-10 |
1,8- -2,0 |
5—7 |
20-30 |
Philadelphus coronarius |
4-5 |
1,5-2,0 |
3- 5 |
15-30 |
Physocarpus opulifolia |
4-5 |
1,5-2,0 |
¦ 1-6 |
20-30 |
Rosa rugosa |
3-4 |
1,0-1,5 |
3—4 |
20 и более |
Sibirea altaiensis |
2—3 |
1,0-1,3 |
2-3 |
5-10 |
|
Поэтому ярусность этого кустарника просматривается четко, но только зимой, когда побеги лишены листьев. У жимолости татарской главная скелетная ось состоит из 5-6 дочерних порядков побегов формирования, у жимолости кавказской, голубой и др. она построена в среднем из 4-6 порядков. Одни стволики более долголетни, другие — менее. Стволик бузины канадской живет не более 3 лет, жимолости камчатской — до 20, У жимолости татарской — 20-25 лет. Долговечность составных скелетных осей — стволиков зависит от длительности основного ц икла. Чем быстрее протекает основной цикл, тем большая часть побега формирования отмирает, тем меньшая входит в состав главной скелетной оси, тем короче эти оси.
В одних случаях становится многолетней большая часть побега формирования, и такие кустарники при небольшом количество ярусов довольно высоки. Например, у жимолости маака высотой 3 м около 4-5 порядков побегов формирования, а у жимолости голубой высотой не более 2 м - до 6 ярусов. В других случаях большая часть побега формирования отмирает, например у бузины красной и бузины черной. Число ССО также различно у разных видов и изменяется в онтогенезе. У Viburnum tinus их 2-3, у V. lantana — 4-6, у жимолости голубой — 3-15 в средневозрастном периоде. У очень многих кустарников других семейств наблюдается та же зависимость: чем короче основные циклы, тем меньшим числом дочерних СПФ образованы основные (главные) скелетные оси и тем интенсивнее, следовательно, проходят циклы развития этих последних (табл. 2). |
Из морфологических признаков систем ССО в первую очередь бросается в глаза степень выраженности ярусов отдельных ветвей. Наиболее четки они у ССО, образованных СПФ второй группы со средней продолжительностью основного цикла. Четкая ярусность возникает за счет того, что СПФ, формирующие стволик, отходят от средних частей СПФ предыдущего порядка в месте их перегиба, в то время как побеги ветвления этой последней продолжают рост и ветвление в горизонтальной плоскости (рис. 7). Если последующие СПФ отходят от предыдущих близ их основания, то ярусы, образуемые разными порядками СПФ, почти сливаются. Если же дочерние СПФ образуются достаточно близко к горизонтально простертой верхушке (материнской), то вся ССО в скором времени, если не найдет опоры, под тяжестью новых систем побегов полегает. Но если побеги ветвления материн– ской СПФ имеют ортотропное направление роста, то ярусов вообще не образуется, откуда бы ни отходили дочерние СПФ.
|

Рис. 7. Побеги формирования у Rosa gallica L . |
На основании всего сказанного можно выделить следующие основные морфологические типы систем ССО: 1) деревцевидные, образованные небольшим числом СПФ, с косовосходящим или ортотропным направлением отдельных ветвей; 2) кустовидные, с ССО небольших размеров вследствие базисимподиального их ветвления; 3) ярусные, с горизонтальным простиранием верхушек СПФ и принадлежащих им СПВ; 4) опирающиеся, лианоидного типа, обвивающие или иным образом удерживающиеся в вертикальном положении с помощью опор, а без таковых полегающие на поверхности почвы; 5) полегающие под тяжестью новых систем побегов, а затем обычно укореняющиеся; 6) приподнимающиеся, образованные немногими генерациями СПФ, приподнимающимися и затем полегающими в основании также под тяжестью вновь образующихся побегов; 7) ползучие, с облигатно простертыми по субстрату побегами формирования и, следовательно, ССО. Характерной особенностью последних трех групп является то, что вторичное утолщение свойственно у них лишь ортотропным частям, плагиотропные же вначале резко снижают вторичный прирост в толщину, а затем и прекращают его вовсе (Колищук, 1968). Особенно ничтожен он у последней группы.
Естественно, каждый стволик ССО обладает у кустарников значительно большей степенью самостоятельности, чем СПФ или СПВ, так как погруженность их оснований в почву или близость с нею дает возможность формировать свои собственные корневые системы. Побег формирования, дающий начало новому стволику (переходящему затем в ССО), может не иметь подземной плагиотропной части [по Л. Е. Гатцук (1970, 1974а), это — ось возобновления, развивающая затем систему моноподиальных побегов оси возобновления] или иметь ее [это — парциальная ось, развивающая систему моноподиальных побегов парциальной оси, по Л. Е. Гатцук, или побег парцилляции, по Ю. Е. Алексееву (1963) ]. Оси возобновления образуют собственно куст, парциальные оси дают начало новому, парциальному кусту. |
В настоящее время главными характеристиками жизненных форм признаются долговечность и количество главных скелетных осей (Серебряков, 1954, 1962). А эти особенности, как мы видели, являются, в свою очередь, функциями основных циклов. Чем быстрее проходит основной цикл (чем он короче), тем менее долговечны главные скелетные оси, тем этих осей больше, растения ниже и кустистее. Именно поэтому основное внимание при описании кустовидной формы роста мы уделили СПФ и основным циклам.