Глава 3. Филогенетические связи в сем. Crassulaceae и статус его восточно-азиатских представителей
на основании сравнения нуклеотидных последовательностей ITS региона ядерной рДНК
Как отмечено нами выше (гл. 1), традиционная система семейства Crassulaceae, основанная на фенотипических признаках и биогеографических данных, существенно противоречит данным молекулярной филогении, и пять из шести подсемейств, установленных А. Berger (1930), являются полифилетическими. Попытка H. 't Hart (1995) преодолеть эту искусственность привела к существенному изменению системы семейства. Автор сократил число подсемейств до двух и выделил несколько триб (см. гл. 1). Одним из таких преобразований было обособление части азиатских представителей Sedoideae sensu А. Berger в подтрибу Telephiinae (Telephium-клада; Hart, 1995; Ham, 1995). Однако как в работах самих авторов, так и в более поздних молекулярно-филогенетических исследованиях монофилитичность этого таксона не была подтверждена, его клада не являлась статистически значимой и демонстрировала явную близость к «Kalanchoe»-кладе (Ham, 1995; Ham, Hart, 1998; Mort et al., 2001, Mayuzumi, Ohba, 2004). А это значит, что некоторые вопросы филогении семейства так и остались неразрешенным (например, полифилитичность рода Sedum s. l.), а кроме того, проявились существенные противоречия морфологических и филогенетических данных, что породило новые проблемы. Так, большинство триб, выделенных H. 't Hart (1995), весьма гетерогенны с точки зрения морфологии слагающих их родов, и для них практически невозможно найти диагностические признаки. Кроме этого, результаты молекулярно-филогенетических анализов поставили под сомнение естественность некоторых родов, выделенных ранее из состава Sedum: Hylotelephium, Orostachys, Rhodiola и Pseudosedum – и считавшихся вполне естественными, что указало на необходимость пересмотра родовых границ восточно-азиатских Crassulaceae. Как уже отмечалось нами ранее, большинство филогенетических построений в семействе основывалось на данных рестриктного анализа и сравнении нуклеотидных последовательностей ДНК хлоропластов, а данные из ядерного генома если и привлекались, то, преимущественно, для анализа взаимоотношений в пределах лишь некоторых родов или группы близких родов (Mes, 1995; Mort et al., 2002). Однако известно, что хлоропластный геном наследуется только по линии одного из родителей, что может существенно влиять на картину филогенетических отношений в группах, где гибридизация играет большую роль в эволюционных процессах. Именно таким таксоном является семейство толстянковых (Uhl, 1976; Spongberg, 1978). В связи с этим, нами была предпринята попытка проанализировать филогенетические отношения в семействе и уточнить положение в нем некоторых восточно-азиатских родов, используя маркер из ядерного генома [последовательности ITS региона рибосомной ДНК (ITS1, 5.8S и ITS2)] и расширенный набор данных. Для этого была создана матрица данных, включающая все известные ITS последовательности толстянковых.
3.1. Филогенетические отношения в семействе Crassulaceae
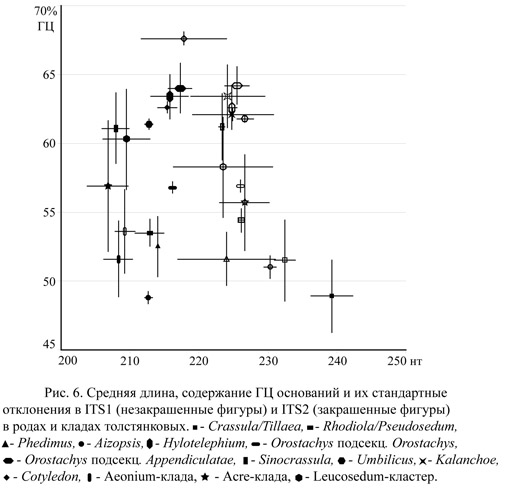
Набор данных включает 272 последовательности представителей семейства. Длина ITS1 варьирует от 202 нт у Aichryson bethencourtianum Bolle до 235 нт у Crassula sieberiana (Schultes) Druce. В большинстве родов длина этого спейсера варьирует незначительно – 223-225 нт, однако у представителей нескольких родов/клад эта величина существенно отличается (рис. 6). Так, у всех членов Aeonium-клады ITS1 короче среднего (209±1,5 нт), а в родах Aizopsis, Crassula и Kalanchoe его длина превышает 230 нт. В пределах родов длина ITS1 изменяется незначительно, однако у видов Cotyledon, Acre- и Kalanchoe клад ее вариабельность высока. Содержание ГЦ оснований также варьирует в пределах набора изученных видов. Некоторые роды (Cotyledon, Hylotelephium, Sinocrassula, Kalanchoe) характеризуются повышенным содержанием ГЦ оснований (>60%; рис. 6), тогда как у других их процент относительно низок (<52%).
Ген 5.8S рДНК почти инвариантен по длине и содержанию ГЦ (50-55,5%) у всех последовательностей за исключением видов Aichryson, которые отличаются синапоморфной делецией нуклеотида на 5' конце гена.
Второй спейсер в целом короче ITS1 [обратная зависимость наблюдается только в роде Crassula, характеризующимся наибольшей длиной ITS2 239±3,0 нт (244 нт у Tillaea aquatica и Crassula mataikona A.P. Druce)], более вариабелен по длине и характеризуется сходным соотношением длины и ГЦ оснований (рис. 6).
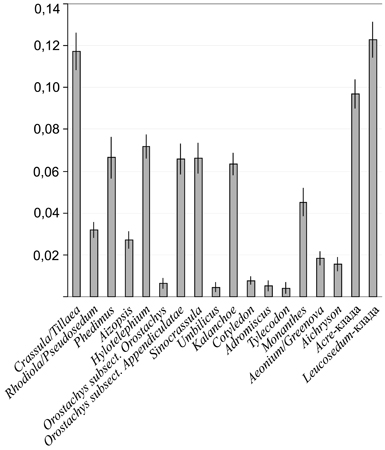
Дивергенция последовательностей спейсерного региона относительно невысока в большинстве родов толстянковых и не превышает 5%. Высоким сходством ITS характеризовались виды Umbilicus, Cotyledon, Adromischus Lemaire, Tylecodon Tolken и родов Aeonium-клады (рис. 7). В то же время в роде Crassula и в Leucosedum-кластере она превышала 12% и была близка к этому значению в Acre-кладе. В целом вариабельность ITS1 выше, чем ITS2 и низка (≤1%) у 5.8S гена.
|
|
|
После того, как было проведено выравнивание последовательностей, из матрицы данных были удалены идентичные, а также неполные последовательности ITS участка. Однако и после этого число видов в матрице оставалось очень высоким, что делало невозможным проведение анализа с использованием сложных эволюционных моделей. Учитывая, что в некоторых группах толстянковых филогенетические взаимоотношения уже анализировались, было решено сократить для них выборки видов, оставив по 2-3 представительных вида (как правило, с наиболее дивергентными последовательностями) на четко выраженную кладу. В конечном итоге матрица данных была сокращена до 188 видов.
Из 591 нуклеотида, включенного в матрицу данных, 156 были инвариантны, а 391 потенциально информативны с точки зрения метода МР. GTR модель эволюции нуклеотидных последовательностей, дополненная гамма-параметром (G) и пропорцией инвариантных позиций (I), была определена как наиболее адекватно описывающая наш набор данных. Её характерной особенностью была повышенная частота двух типов нуклеотидных замен, Ц↔T (4.4802) и A↔Г (2.7161).
Рис. 7. Дивергенция последовательностей (р-дистанции) и их стандартное отклонение в родах и кладах толстянковых.
|
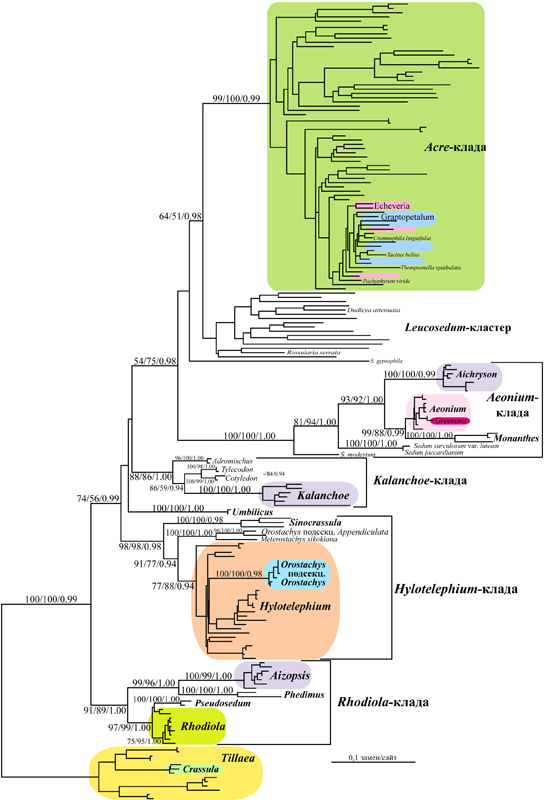
Рис. 8. Филогенетическое дерево сем. Crassulaceae, основанное на сравнении 188 ITS рДНК последовательностей (591 нуклеотид) методом максимального правдоподобия (GTR+I+G модель). Цифрами указана устойчивость основных ветвей, рассчитанная для NJ/MP/BI-анализов методом бутстрепа (NJ и MP) и апостериорных вероятностей (BI).
|
При анализе этой матрицы методом ML было получено древо, представленное на рис. 8. На нем виды толстянковых сгруппированы в 6 основных устойчивых клад (поддержанных значениями бутстрепа ≥50% и апостериорных вероятностей ≥0.90). Это клады подсемейства Crassuloideae (слагаемого родами Crassula и Tillaea), «Kalanchoe» (роды Adromischus, Tylecodon, Cotyledon и Kalanchoe; >85% бутстрепа), «Hylotelephium» (роды Hylotelephium, Orostachys и Sinocrassula), «Rhodiola» (роды Rhodiola, Pseudosedum, Phedimus и Aizopsis) и монородовая клада «Umbilicus». Виды рода Sedum s. str. на древе распределены между тремя кладами, занимающими вершинное положение (рис. 8), три вида (S. modestum Ball и пара S. jaccardianum Maire et Wilczek/S. surculosum Cosson) располагаются в парафилетической последовательности в основании высоко устойчивой (100 % ВР, 1.00 РР) «Aeonium»-клады, слагаемой родами Monanthes Haworth, Aichryson Webb et Berthelot и полифилетичной Aeonium/Greenova-кладой. Большинство видов Sedum входят в состав парафилетичной группы «Leucosedum» или полиродовой «Acre»-клады. Еще один вид (S. gypsophillum Turner) является сестринским таксоном этой вершинной группы. Большинство клад, упомянутых выше, имеют высокую бутстреп-поддержку (>85%) и значения апостериорных вероятностей (>0.95), однако порядок их ветвления в нашем анализе установить не удалось. Из 28 традиционных родов толстянковых, проанализированных нами, монофилия была подтверждена только для 10. Виды Rosularia, Dudlea, Graptopetalum Rose, Tacitus Moran et Meyrán, Cremnophila Rose, Echeveria и Pachyphytum Link, Klotzsch et Otto вошли в состав «Leucosedum»-кластера или «Acre»-клады, где они занимали вершинное положение среди видов Sedum. Помимо этого нами установлено, что клады Hylotelephium/Orostachys, Rhodiola/Pseudosedum и Greenova/Aeonium перекрывают родовые границы (рис. 8).
Анализ большого числа ITS последовательностей толстянковых позволил нам построить первую филогению семейства, основанную на ядерном геноме, и сравнить ее с другими топологиями, основанными на последовательностях ДНК хлоропластов: протеин-кодирующем гене matK (Mort et al., 2001) и trnL-trnF межгенном спейсере (Mayusumi, Ohba, 2004). Необходимо отметить, что в целом все три использованных маркера устанавливают в сем. Crassulaceae одни и те же основные клады со сравнимой статистической значимостью и сходным образом отражают филогенетические взаимоотношения между ними. Сходство нескольких топологий, базирующихся на информации из разных участков генома, имеющих собственную эволюционную динамику, позволяет предположить, что филогенетические гипотезы, следующие из этих построений, достоверно отражают эволюционные взаимоотношения в семействе.
Высокая статистическая значимость клад разного уровня нашего древа также позволяет говорить, что ITS является хорошим маркером для реконструкции филогении семейства на разных уровнях, от видового в некоторых родах, до семейственного. Следует заметить, что роды толстянковых отличаются скоростями накопления мутаций в ITS. Так, в большинстве родов макаронезийской группы, Rhodiola и некоторых других различия между видами слабо выражены, тогда как в родах Hylotelephium, Sedum и Kalanchoe они существенны и позволили с высокой степенью достоверности установить родственные связи в этих таксонах.
|
Наибольшую информативность ITS последовательности имеют на уровне род-триба. Во всех основных кладах взаимоотношения между родами имеют высокую статистическую значимость и полностью соответствуют данным, полученным с использованием хлоропластных маркеров. Однако порядок ветвления основных клад древа напротив, имеет слабую статистическую значимость, и, вероятно, применение этого маркера для установления взаимоотношений между ними малоперспективно.
Данные, полученные нами, еще раз подтвердили существование противоречий между традиционной таксономией семейства, основанной на фенотипических признаках, и филогенетическими отношениями между видами и родами, установленными в предшествующих анализах. Согласно молекулярным данным, семейство слагается как минимум 7 кладами (Ham, 1995; Mort et al., 2001; Mayuzumi, Ohba, 2004), из которых только одна соответствует подсемейству Crassuloideae в понимании Berger. Другие же клады, объединяющие в своем составе представителей нескольких традиционных подсемейств, в системе H. 't Hart (1995) имеют ранг триб или подтриб (Kalanchoeae и Sedeae; Telephiinae и Sedinae). Следует отметить, что подтриба Sedinae слагается четырьмя кладами, не имеющими формального таксономического статуса: «Acre», «Leucosedum», «Sempervivum» и «Aeonium». Наш анализ подтвердил монофилию 5 из 7 клад, полученных предшествующими исследователями, еще одна («Leucosedum») была установлена только топологически. А роды, слагающие «Telephium-кладу» sensu H. 't Hart (1995), образовали три независимые ветви на древе: «Rhodiola», «Hylotelephium» и монородовая «Umbilicus». Установленная нами топология в значительной мере соответствует системе семейства, предложенной H. t'Hart (1995), но положение трибы Kalanchoeae на нашем древе нарушает монофилию трибы Sedeae. Согласно построениям H. t' Hart, поддержанным данными анализа хлоропластных маркеров (Mort et al., 2001; Mayuzumi, Ohba, 2004), Kalanchoeae является базовой в подсемействе Sedoideae, а на нашем древе она располагается вслед за тремя ветвями, относимыми к Telephiinae (рис. 8), хотя и с невысокой статистической значимостью. Следует отметить, что ни одна, ни другая филогенетическая гипотеза не подтверждаются фенотипическими и/или биогеографическими данными. J.T. Baldwin (1938) полагал, что Kalanchoe и близкие к нему роды ведут происхождение от предковой формы, близкой к Crassula, тогда как C.H. Uhl (1948) отвергал вероятность подобного сценария. Преимущественное распространение членов «Kalanchoe»-клады и «Crassula» в Южной Африке позволяют предположить их возможное родство, однако почти космополитное распространение Tillaea, еще одного представителя «Crassula»-клады, дает возможность для построения других гипотез.
Топология филогенетического древа толстянковых, основанная на сравнении я-р ДНК (настоящее исследование) ставит под сомнение правомерность выделения в составе подсемейства Sedoideae двух триб.
Хотя «Telephium»-клада и не была достоверно установлена ни в одном из трех анализов (Mort et al., 2001; Mayuzumi, Ohba, 2004; настоящее исследование), этот факт нельзя рассматривать как доказательство отсутствия родства между членами клады (трибы).
История «Telephium»-клады (в настоящее время имеющей формальный статус подтрибы) берет начало с первых молекулярных исследований, использовавших результаты рестрикционного анализа хлоропластной ДНК (Ham, 1995). Изначально эта группировка была установлена с очень низкой (25%) статистической значимостью, но в то время этому факту не придавали значения. Исследователи решили, что общность современного распространения большинства членов клады является доказательством их происхождения от общего предка.
В более позднем анализе хлоропластного matK гена клада вновь не была установлена (Mort et al., 2001), однако в этом гене было обнаружено несколько предположительно синапоморфных инделей, отличающих роды, относимые к ней, от других толстянковых, что также было истолковано как подтверждение общности их происхождения. Следует заметить, что в то же время были обнаружены и индели, свойственные только Crassula и Umbilicus, однако их посчитали результатом гомоплазии (Mort et al., 2001).
Фенотипическими признаками, объединяющими членов клады, по мнению H. t' Hart (1995) являются пятичленные цветки с обдиплостемонным андроцеем и свободными лепестками, как правило, плоские зубчатые или пильчатые листья и т.д. Однако ни один из этих признаков не является уникальным для совокупности членов трибы (Mayuzumi, Ohba, 2004). Как справедливо отметили S. Mayuzumi и H. Ohba (2004), не существует фенотипических признаков, однозначно характеризующих членов этой клады и подтрибы, что, впрочем, справедливо и для большинства других «новых» внутрисемейственных таксонов Crassulaceae.
Помимо восточно-азиатских родов к подтрибе Telephiinae относят и род Umbilicus, традиционно включаемый в состав Cotyledonoideae и распространенный преимущественно в средиземноморском регионе. Его ареал практически не пересекается с ареалами других членов подтрибы (исключение – Phedimus), а морфология существенно отличается от морфологии представителей других родов Telephiinae. Родственные связи Umbilicus долгое время оставались не раскрыты, однако в анализе ITS последовательностей S. Mayuzumi и H. Ohba (2004) были установлены сестринские отношения между ним и кладами «Rhodiola» и «Phedimus», эквивалентной нашей «Rhodiola»-кладе, но с невысокой статистической достоверностью. Наш анализ, базирующийся на существенно большем числе последовательностей и опирающийся на информацию о вторичной структуре спейсеров, отвергает сестринские отношения этих групп, и демонстрирует, что Umbilicus достоверно является независимой ветвью (56 и 74 ВР, 0,99 РР поддержка для ветви, отделяющей «Rhodiola»-кладу и Umbilicus). Морфологические признаки (пятимерный цветок со сросшимися венчиком) сближают Umbilicus с членами Cotyledonoideae (Berger, 1930), в настоящее время относимыми к трибе Kalanchoeae, но основное хромосомное число (х=24), морфология хромосом и преимущественно средиземноморское распространение выделяют этот род из прочих Cotyledonoideae (Uhl, 1948). Однако эти же признаки отличают Umbilicus и от членов Telephium-клады (Telephiinae). Таким образом, вопрос о положении рода Umbilicus в семействе так и остается не решенным. Молекулярные данные подтвердили обособленность Umbilicus, но не смогли определить его родство.
Независимо от положения рода Umbilicus в подтрибе Telephiinae, мы можем заключить, что клады «Rhodiola» и «Hylotelephium» проявляют мало близости друг к другу в молекулярных филогениях (Mort et al., 2001; Mayuzumi, Ohba, 2004, настоящее исследование). Такой результат можно было бы объяснить гомоплазией последовательностей, но его повторение независимо в трех анализах (хлоропластного гена matK и trnL-trnF межгенном спейсере (Mort et al., 2001; Mayuzumi, Ohba, 2004) и я-рДНК, настоящее исследование), делает это объяснение маловероятным. Более достоверным нам кажется то, что клады если и ведут происхождение от общего предка, то их дивергенция произошла очень давно. В пользу этого предположения говорит то, что представители клад имеют различное распространение, что, возможно, отражает их независимые истории. Аркто-альпийский род Rhodiola, слагающий ядро одноименной клады, приурочен преимущественно к высокогорьям центральной и Юго-Восточной Азии и включает ограниченное число широко распространенных видов (R. rosea, R. integrifolia), Phedimus представляет западную ветвь этой клады, и только современный ареал Aizopsis в значительной мере пересекается с ареалами Hylotelephium и Orostachys, типичных восточно-азиатских родов (элементов). Принимая гипотезу южно-африканского или средиземноморского происхождения семейства (Ham, Hart, 1998), мы однозначно принимаем последующую миграцию предковых форм в Азию и их последующую их диверсификацию здесь. Однако вопрос о том, случилось это один (монофилетичная «Telephium»-клада) или несколько раз (несколько независимых клад), пока остается не решенным.
Более ясным представляется нам статус и структура вершинной группы древа, слагаемой видами рода Sedum s. str. и представителями традиционных подсемейств Sempervivoideae и Echeverioideae (рис. 8). Хотя эта клада и объединяет представителей по крайней мере 3 подсемейств А. Berger, она полностью соответствует подтрибе Sedinae sensu H. 't Hart (1995). В соответствии с системой H. ‘t Hart, подтриба слагается четырьмя группами: устойчивыми макаронезийской, «Sempervivum» (не анализировалась в настоящем исследовании), «Acre» кладами и парафилетической «Leucosedum». Виды Sedum входят в состав всех этих клад, демонстрируя полифелитичность рода. Предположения об его сборной природе высказывались неоднократно (Ham, 1995; Mayzumi, Ohba, 2004; Ohba, Mayzumi, 2004 и др.), сделано множество попыток выделить из состава Sedum монофилетические группы и таким образом преодолеть эту искусственность (Ohba, 1977, 1995; Grulich, 1984; и др.), однако решение пока не найдено.
3.2. Филогенетические взаимоотношения в родах восточно-азиатских Crassulaceae
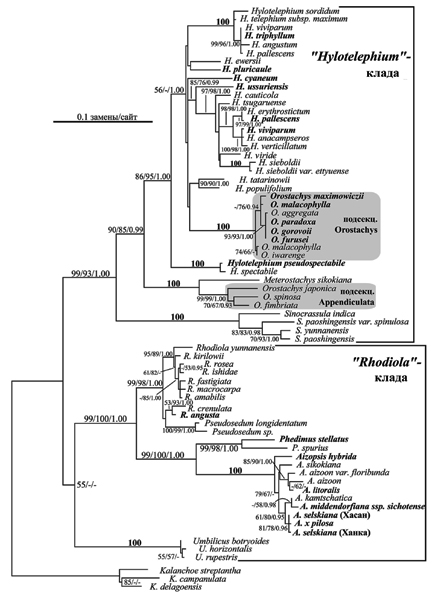
Рис. 9. Филогенетическое древо представителей подсем. Sedoideae, основанное на сравнении 69 ITS рДНК последовательностей методом максимального правдоподобия (GTR+I+G модель) (по: Гончарова и др., 2006). Цифрами указана устойчивость ветвей, рассчитанная для NJ/MP/BI-анализов (100=100/100/1.00). Жирным шрифтом отмечены виды, последовательности которых определены в Гончарова и др. (2006). Серым цветом выделены две независимые клады видов Orostachys.
|
Согласно результатам проведенного нами общего анализа семейства, восточно-азиатские роды толстянковых, относимые к подсемейству Sedoideae (Berger, 1930) или Telephiinae (Hart, 1995), вероятно, не образуют монофилетической клады, а формируют три независимые ветви: «Rhodiola»- и «Hylotelephium»-клады и «Umbilicus» (рис. 8). Также было показано, что эти клады проявляют родство к «Kalanchoe»-кладе. Поскольку в общем анализе из матрицы данных был удален ряд дальневосточных видов, нами был проведен еще один анализ, в котором упор был сделан на выяснение взаимоотношений только в кладах «Rhodiola» и «Hylotelephium» [рис. 9] (Гончарова и др., 2006).
Для этого в меньшую матрицу данных были включены все различающиеся последовательности из этих родов, а также Umbilicus, Sinocrassula, Meterostachys и Orostachys (всего 69 последовательностей). Как и в предыдущем анализе GTR модель эволюции нуклеотидных последовательностей, дополненная гамма-параметром (G) и пропорцией инвариантных позиций (I) и имеющая сходные параметры, была определена как наиболее адекватно описывающая этот набор данных. Из 604 нт, использовавшихся для анализа, почти половина (295 нт) были информативными с точки зрения метода MP, 251 – инвариантными и 58 – неинформативными.
Результаты ML анализа отображены на рис. 9. Как и в общем анализе, на древе последовательности дальневосточных представителей подсемейства сгруппированы в две клады, условно названные нами «Hylotelephium» и «Rhodiola». В первой хорошо поддержанной (>93 BP, 1.00 PP) кладе род Sinocrassula образует базальную ветвь, за ним следует устойчивая (100 ВР) сестринская пара Meterostachys-Orostachys подсекц. Appendiculata (Boriss.) H.Ohba, а клада видов Hylotelephium, достоверно (>85% BP, 1.00 PP) включающая представителей Orostachys подсекц. Orostachys, является вершинной.
В отличие от топологии, представленной на рис. 8, принадлежность базального рода Umbilicus к группе Rhodiola поддержана низким значением бутстрепа только в NJ анализе. За ним следуют две устойчивые (99-100% ВР, 1.00 РР) клады, каждая из которых объединяет представителей двух родов: пара Phedimus-Aizopsis и клада видов Rhodiola, включающая два вида Pseudosedum. Данная топология характеризует только 4 из 8 проанализированных родов как монофилетические: Phedimus, Aizopsis, Umbilicus и Sinocrassula, а их клады поддержаны высокими значениями бутстрепа (98-100%) и апостериорных вероятностей (1.00).
|
Дивергенция между названными кладами достаточно хорошо выражена, и все они характеризуются длинными индивидуальными ветвями. Различия между последовательностями в кладах не столь существенны, однако они позволили с высокой степенью достоверности определить порядок взаимоотношения между большинством видов. Наиболее высокой дивергенцией отличаются представители Sinocrassula, Phedimus и Hylotelephium, а в родах Rhodiola и Aizopsis напротив, различия между видами невелики (рис. 9).
Сравнивая длины ветвей в двух установленных кладах рода Orostachys, можно заметить, что различия между видами подсекц. Appendiculata более выражены, и их последовательности четко отличаются друг от друга (25-35 замен), тогда как в подсекц. Orostachys различия не превышают 2-4 нт. Эти клады различаются не только заменами нуклеотидов, но и наличием специфических инделей (рис. 10), не учитывавшихся при филогенетическом анализе. Наибольшее их количество обнаружено в более вариабельном ITS1. Примечательно, что клады подсекций практически не отличаются длиной этого спейсера и всего спейсерного региона, т.е. делеции в одних участках ITS компенсируются вставками в других.
На филогенетическом древе некоторые виды (Hylotelephium viviparum, H. рallescens, Aizopsis selskianum и Orostachys malacophylla) представлены двумя последовательностями. При этом только в случае A. selskianum не обнаружено различий между ними. Экземпляры O. malacophylla отличаются 4 заменами, а различия между образцами Hylotelephium viviparum, H. рallescens еще более существенны – более 50 замен и инделей. Не удивительно, что эти последовательности располагаются в разных частях клады рода (рис. 9). Вероятнее всего такие различия обусловлены расхождениями в понимании объема вида либо в неверном его определении.
При общей высокой дивергенции между видами Hylotelephium в роде выделяется несколько устойчивых групп видов, члены которых характеризуются высоким сходством последовательностей ITS (≤10 замен). Это пара H. erythrostictum/ H. pallescens, клада H. verticillatum/ H. anacampseros/ H. viviparum и устойчивая H. sordidum-клада, объединяющая 6 последовательностей (рис. 9).
Сравнение нуклеотидных последовательностей внутреннего транскрибируемого спейсера рибосомной ДНК 69 представителей 10 родов подсем. Sedoideae, имеющих преимущественно азиатское распространение, позволило установить их филогенетические связи. Полученные результаты в значительной мере согласуются с данными предшествующих исследований, основанных на меньших наборах данных и других маркерах (Mayuzumi, Ohba, 2004). Основной упор был сделан нами на анализе взаимоотношений между представителями родов Orostachys и Hylotelephium, монофилия которых была поставлена под сомнение (Mayuzumi, Ohba, 2004), а также родов Phedimus и Aizopsis, рассматриваемых некоторыми авторами как синонимы (Eggli, 1995; Ohba et al., 2000).
Orostachys считается одним из наиболее четко отграниченных морфологически членов подсемейства (Ohba, 1978). Этот род характеризуется полурозеточными жизненными формами и терминальными колосовидными (початковидными) соцветиями. В роде выделяют две секции: Orostachys и Schoenlandia (Ohba, 1978), последняя принимается некоторыми авторами как подрод Schoenlandia (H. Ohba) Byalt (Бялт, 1999а, 1999б) или самостоятельный род Kungia K.T. Fu (Fu, Ohba, 2001) и не представлена на российском Дальнем Востоке. В свою очередь, виды типовой секции классифицируют в двух подсекциях: Orostachys и Appendiculatae, – различающихся формой листа (плоские или вальковатые) и наличием или отсутствием на них придатков (шипов, хрящеватых выростов и др.). Молекулярно-филогенетические данные подтвердили близость видов в каждой подсекции, но установили отсутствие родства между подсекциями. Более того, типовая подсекция Orostachys была достоверно (>85% BP, 1. 00 РР; рис. 9) помещена среди членов рода Hylotelephium. Эти взаимоотношения трудно обосновать с точки зрения морфологии, однако конгруэнтность филогений, основанных на маркерах из ядерного (настоящее исследование) и хлоропластного (Mаyuzumi, Ohba, 2004) геномов, еще раз подтверждает их достоверность. Общими признаками для Hylotelephium и Orostachys подсекц. Orostachys является морфология листа (плоские, без отростков) и гинецея, но полурозеточные биоморфы, дициклические монокарпические побеги и колосовидные соцветия отличают последний таксон.
Виды Orostachys с вальковатыми листьями, имеющими отростки на концах (подсекц. Appendiculatae), также сгруппированы в устойчивую кладу, и можно предположить, что данная морфологическая синапоморфия имеет большее таксономическое значение, чем это предполагалось ранее. Этот вывод подтверждает и родство Orostachys подсекц. Appendiculatae и монотипного рода Meterostachys (рис. 9), также характеризующегося вальковатыми листьями с шипами.
Очевидно, что виды родов Hylotelephium, Orostachys и Meterostachys близки между собой (рис. 9) и ведут происхождение от общего предка. Однако структура их общей клады противоречит традиционным объемам родов и предполагает необходимость таксономических преобразований. Так, признание монофилии клады Hylotelephium-Orostachys подсекц. Orostachys, следующее из результатов молекулярно-филогенетических анализов (Mаyuzumi, Ohba, 2004; настоящее исследование), подразумевает, что все члены клады должны быть включены в род Orostachys, пользующийся правом приоритета (International code…, 2000). При этом существующий диагноз рода должен быть изменен, для того чтобы охватить морфологическое разнообразие нового комбинированного рода. В настоящее время это вряд ли возможно, поскольку не известны признаки, являющиеся общими для всех видов рода и отличающие их от видов, не входящих в его состав.
|
Согласно представленной топологии, Orostachys подсекц. Appendiculatae заслуживает выделения в самостоятельный род. Вопрос принадлежности Meterostachys sikokiana Nakai к этому таксону остается открытым. Сестринские отношения между двумя таксонами не противоречат как самостоятельности последнего, так и возможности их объединения. Решение этого вопроса зависит от веса, придаваемого признакам, общим для представителей клады и отличающим их от других ветвей. На наш взгляд, Meterostachys довольно четко отличается от членов подсекц. Appendiculatae облигатно розеточной биоморфой, пазушными метельчатыми соцветиями и числом хромосом (16 и 12 соответственно; Uhl, Moran, 1972). Кроме этого, на молекулярном уровне члены Orostachys подсекц. Appendiculatae характеризуются аутапоморфными инделями (рис. 10).
Рис. 10. Примеры синапоморфных инделей и замен (выделены серым цветом) в ITS1, отличающих подсекции рода Orostachys. Элементы вторичной структуры спейсера (1 -стебель 1, 2 – стебель 3) показаны в соответствии с моделью, разработанной для ITS1 Rhodiola rosea. Толстой линией отмечены коадаптивные замены в стебле 1. Штриховой линией показаны индели.
|
Топология представленного нами филогенетического древа ставит вопрос о самостоятельности еще двух таксонов: родов или подродов Phedimus и Aizopsis. До недавних пор они рассматривались как секции или подроды в составе Sedum (Berger, 1930; Ohba, 1978), однако позже были выделены в самостоятельные роды (Grulich, 1984; Ohba, 1995; Бялт, 2001). Уже первые молекулярно-филогенетические анализы показали близость этих таксонов (каждый был представлен одним видом), что и послужило основанием для их объединения в род Phedimus (Hart, 1995). На нашем древе Phedimus и Aizopsis также являются сестринскими кладами, каждая из которых включает типовой вид соответствующего рода (P. stellatus и A. aizoon соответственно; рис. 7). Данная топология не отрицает как самостоятельность этих родов, так и возможность их объединения. Мы считаем, что различия между членами клад в окраске (розовые или белые у Phedimus и желтые у Aizopsis) и морфологии цветков, распространении (преимущественно европейское у Phedimus и азиатское у Aizopsis) являются существенными и отличают их как самостоятельные роды.
В нашем анализе представлено большинство видов рода Hylotelephium, что позволяет провести оценку существующей внутриродовой системы. В составе этого таксона выделяют две секции и две подсекции, различающиеся жизненными формами и положением побегов в пространстве. Описывая эти таксоны, H. Ohba (1978) придавал большое значение сходству морфологии слагающих их видов и предполагал, что оно отражает общность их происхождения. Однако молекулярно-филогенетические данные характеризуют все эти подроды и секции как искусственные и предполагают независимое возникновение признаков, характеризующих их. В составе Hylotelephium выделяются устойчивые клады, однако в настоящее время трудно назвать морфологические признаки, объединяющие их членов. Несоответствие традиционной внутриродовой классификации и данных молекулярно-филогенетических исследований отмечено и для рода Rhodiola (Mаyuzumi, Ohba, 2004).
Таким образом, наше исследование подтвердило наличие существенных противоречий между традиционными взглядами на эволюцию и родственные связи восточно-азиатских очитковых. Очевидно, что необходимы дальнейшие исследования для того, чтобы достоверно установить таксономическую структуру подсемейства и естественные границы родов, а также положение этих таксонов в семействе.
Оригинальный рисунок А.Р. De Candolle www.illustratedgarden.org/mobot/rarebooks, Rhodiola pinnatifida nature.chita.ru.
Читать далее
|