Глава 5
Особенности распределения эпифитных лишайников на дубе монгольском и дубе зубчатом
Как видно из приведенного в предыдущей главе аннотированного списка, на юге Приморского края флора лишайников-эпифитов, встречающихся на дубе монгольском и дубе зубчатом, насчитывает 201 вид, относящихся к 61 роду и 26 семействам. Из них 197 видов (60 родов и 25 семейств) обнаружено на дубе монгольском, соответственно 82 вида (33 рода и 13 семейств) - на дубе зубчатом. Общими для дуба монгольского и дуба зубчатого являются 78 видов.
Несмотря на сходные экологические и климатические условия, в которых дуб монгольский и дуб зубчатый формируют сообщества на юге Приморья, состав стволовых эпифитных лишайниковых группировок на разных видах дуба существенно различается.
Распределение видов лишайников-доминантов оказалось неравномерным в пределах геоботанических площадей, заложенных в Хасанском районе. На разных лихенометрических площадках в пределах одной пробной площади доминируют разные виды. Вид, доминирующий на одной площадке, часто является малообильным на других. Рядом стоящие деревья имеют неодинаковый состав доминирующих видов. Такая ситуация возможна только при жесткой конкуренции между особями и видами лишайников, когда решающее воздействие на характер формируемого лишайникового сообщества оказывает сам факт первопоселения.
Доминантом становится тот, кто поселяется первым и больше имеет диазачатков. Это позволяет говорить о существенной роли ценотических факторов (конкуренция, аллелопатия, пространственное исключение) при формировании локальных лихеногрупп. В связи с тем, что особи лишайников живут долго, очень медленно растут и не образуют многоярусных сообществ, факт первопоселения при формировании локальных лихеногрупп играет более важную роль, чем в сообществах семенных растений (Работнов, 1978). Заселившись однажды, лишайники «не отдают» свою экологическую нишу без резкого воздействия внешних экологических факторов.
На дубе зубчатом доминируют виды из родов Heterodermia, Phaeophyscia, Physconia, реже, чем на дубе монгольском, встречаются представители Caloplaca, Pyxine, Schismatomma. Совсем редко на дубе зубчатом встречаются Lecidella elaeochroma и виды из рода Pertusaria, а Tephromela atra не обнаружен ни разу. Leptogium burnetiae и Collema subnigrescens встречены на дубе зубчатом в небольшом количестве на самом юге Хасанского района, а на дубе монгольском эти виды обнаружены севернее, где дуб зубчатый встречается реже.
На дубе монгольском доминируют виды из родов Caloplaca, Heterodermia, Myelochroa, Lecidella, Pyxine, Schismatomma, изредка на отдельных стволах дуба монгольского доминирующим видом является Phaeophyscia hispidula. В то же время на других деревьях в пределах массива этот вид вообще отсутствует. Более часто, чем на дубе зубчатом, на дубе монгольском встречаются Anaptychia isidiata, Heterodermia hypoleuca, Pertusaria sp., Phaeophyscia hispidula. Tephromela atra обнаружен только на дубе монгольском.
В процессе работы по выявлению видового состава на дубе зубчатом и дубе монгольском в дубняках Хасанского района, обнаружилось, что некоторые виды лишайников являются взаимозамещающими на разных видах дуба - это экологически викарирующие виды лишайников.
На разных видах дуба доминируют разные виды некоторых родов лишайников. Так, например, на дубе зубчатом из рода Heterodermia доминирует H. diademata, а на дубе монгольском его вытесняет полностью H. hypoleuca, на дубе зубчатом он представлен очень скудно и не в таком хорошем состоянии, как на дубе монгольском. Из рода Phaeophyscia на дубе зубчатом абсолютно доминирует среди эпифитных лишайников P. hirtuosa, а на дубе монгольском этот вид встречен очень локально - только на двух стволах. Вместо него из этого же рода на дубе монгольском присутствует P. hispidula, причем обильнее, чем на дубе зубчатом. Как уже отмечено выше, на отдельных стволах дуба монгольского Phaeophyscia hispidula абсолютно доминирует над остальными эпифитными лишайниками.
Род Pyxine представлен двумя видами - P. sibirica и P. sorediifera. На обоих видах дуба они являются необильными видами, хотя на дубе монгольском оба встречаются чаще и являются более обильными. Однако на дубе зубчатом P. sibirica встречается намного чаще, чем P. sorediata (обнаружен лишь на одной площадке), в то время как на дубе монгльском P. sorediata встречается в 2,5 раза чаще, чем P. sibirica. Род Physconia представлен четырьмя видами - Physconia detersa, P. subpulverulenta, P. kurokawae, P. lobulifera, из которых P. detersa, P. subpulverulenta, P. kurokawae являются доминирующими и обильными на дубе зубчатом, а на дубе монгольском очень редко встречается только Physconia detersa.
Можно предположить, что различный состав лишайниковых сообществ зависит от структуры коры - более грубой и неровной на поверхности у дуба зубчатого, чем у дуба монгольского. Кроме того, величина рН коры влияет на доступность питания в стекающей по столам воде (Бязров, 2002), а различия в питании определяют доминирование и обилие тех или иных видов лишайников. Изменения в химизме и рН коры деревьев могут вносить регулярно проходящие низовые пожары в дубовых лесах Хасанского района.
Лишайники различных жизненных форм распределяются на дубе монгольском и дубе зубчатом по-разному (таблица 2). На стволах обоих видов дуба доминирующими среди других жизненных форм являются листоватые лишайники, которые произрастают на различной высоте от почвы: от десятков сантиметров до нескольких метров (ограничением может быть только молодой возраст верхних ветвей). Среднее проективное покрытие листоватых лишайников на дубе монгольском составляет 28%, а на дубе зубчатом - 19%. Максимальное проективное покрытие листоватых лишайников на площадках на дубе монгольском составляет 71%, а на дубе зубчатом - 75%. Видовой состав этой жизненной формы неодинаков на дубе монгольском и дубе зубчатом.
Таблица 2.
Проективное покрытие эпифитных лишайников разных жизненных форм на двух видах дуба
|
Дуб монгольский |
Дуб зубчатый |
Жизненная
форма |
Среднее |
Максимальное |
Среднее |
Максимальное |
Листоватые |
28% |
71% |
19% |
75% |
Накипные |
14% |
66% |
8% |
38% |
Кустистые |
Не редко встречаются |
В массе молодые зачатки |
Редко встречаются на высоте более 2,5 м |
Молодые зачатки не встречены |
Взрослые кустистые лишайники очень редко встречаются на дубе зубчатом, и только на высоте более 2- 2,5 метров от почвы, а молодые зачатки этой жизненной формы практически не встречаются вообще. В то же время на дубе монгольском кустистые лишайники довольно часто и местами обильно встречаются на высоте от 0,5 м до нескольких метров, а их молодые зачатки иногда бывают очень многочисленны. Интересен и тот факт, что кустистые эпифитные лишайники здесь представлены только родом Ramalina.
Среднее проективное покрытие накипных лишайников на дубе монгольском составляет 14%, а на дубе зубчатом - 8%. Максимальное проективное покрытие накипных лишайников на дубе монгольском составляет 66%, а на дубе зубчатом - 38%. Доминирующими видами из накипных лишайников на дубе монгольском являются Caloplaca gordejevi, Schismatomma pericleum, Lecanora sp., Lecidella elaeochroma.
В результате индикаторного анализа (TWI NSPAN), в котором учитывались проективное покрытие и встречаемость каждого вида, выделено пять групп лихенометрических площадок на дубе монгольском в Хасанском районе (пробная площадь З-1) (табл. 3), в которых доминируют следующие виды: Lecidella elaeochroma – в 1-й группе, Myelochroa entotheiochroa - в 2-й группе, Pyxine sibirica - в 3-й группе, Phaeophyscia hirtuosa - в 4-й группе, Flavoparmelia caperata - в 5-й группе. Каждая из этих пяти групп характеризуется конкретными видами лишайников, отличающими ее от других групп. Эти виды имеют наибольшую встречаемость и проективное покрытие на площадках данной группы (показано цифрами в таблице 3).
Группа 1, кроме Lecidella elaeochroma, характеризуется видами Caloplaca gordejevi, Myelochroa subaurulenta, Parmotrema chinense, Pyxine sorediata. Группа 2, помимо доминантного вида Myelochroa entotheiochroa, включает Schismatomma pericleum.
В группе 3 вместе с Pyxine sibirica также наблюдается Punctelia borreri. В группе 4 особенности исчерпываются доминированием Phaeophyscia hirtuosa, а в группе 5 к Flavoparmelia caperata присоединяется Ochrolechia parella. Наличие таких групп видов, возможно, связано с наличием определенных типов лишайниковых сообществ, каждое из которых контролируется своим доминантом.
Отношение выделенных групп к высоте над уровнем почвы показано на рис. 2.
Тaблица 3.
Группы лихенометрических площадок на дубе монгольском, выделенные с помощью индикаторного анализа (TWINSPAN), с учетом проективного покрытия лишайников
Группа 1
Caloplaca gordejevi |
82 |
67 |
66 |
60 |
60 |
Lecidella elaeochroma |
88 |
58 |
25 |
33 |
40 |
Myelochroa subaurulenta |
52 |
|
8 |
6 |
|
Parmotrema chinense |
70 |
19 |
50 |
13 |
40 |
Pyxine sorediata |
52 |
12 |
16 |
|
|
Группа 2
Myelochroa entoteiochroa |
17 |
74 |
83 |
80 |
60 |
Schismatomma pericleum |
17 |
48 |
|
6 |
|
Группа 3
Punctelia borreri |
17 |
9 |
41 |
|
20 |
Pyxine sibirica |
|
3 |
58 |
|
40 |
Группа 4
Группа 5
Flavoparmelia caperata |
17 |
|
16 |
|
100 |
Ochrolechia parella |
11 |
48 |
8 |
13 |
60 |
Cуществует слабая связь между высотой площадки над уровнем почвы и выделенными группами видов. Группы видов, выделенные методом TWINSPAN, оказались слабо связанными с экспозицией. Так, например, площадки, отнесенные к первой группе, распространены на южной, западной, юго-западной, восточной и северо-западной экспозициях, на высоте от 0,6 до 1,6 м от поверхности почвы; вторая группа площадок - на западной, юго-западной, юго-восточной, восточной и северо-западной экспозициях, на высоте от 0,3 до 1,7 м; третья группа - на юго-восточной, северо-восточной, западной и северо-западной экспозициях, на высоте от 0,5 до 1,5 м; четвертая группа - на юго-восточных, восточных, северо-западных, западных и юго-западных экспозиции, на высоте от 0,5 до 1,6 м; пятая группа - на юго-западной, западной и юго-восточной экспозициях, на высоте от 1 до 1,5 м.
Методом дисперсионного анализа выявлялась зависимость проективного покрытия от экспозиции. На рис. 3, показано, что для разных жизненных форм лишайников эта зависимость различна. Проективное покрытие листоватых лишайников увеличивается на северо-западных и северо-восточных экспозициях, а на южных, юго-восточных, восточных экспозициях уменьшается. Проективное покрытие кустистых лишайников с изменением экспозиции практически не изменяется, а у накипных лишайников явно увеличивается на южных, юго-западных и юго-восточных экспозициях.
Так же неоднозначно происходит изменение проективного покрытия лишайников разных жизненных форм в зависимости от высоты над уровнем почвы (рис.4). Проективное покрытие листоватых лишайников слабо изменяется с высотой, но заметна тенденция к его снижению с увеличением высоты. Проективное покрытие накипных лишайников, по сравнению с листоватыми, более четко уменьшается с высотой. Кустистые лишайники на Quercus mongolica имеют низкую встречаемость вообще и низкое проективное покрытие, которое не зависит от высоты над уровнем почвы.
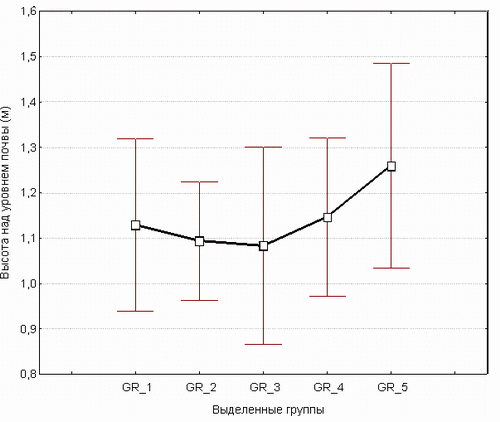
Рис. 2. Отношение выделенных групп видов лишайников к высоте над уровнем почвы
|
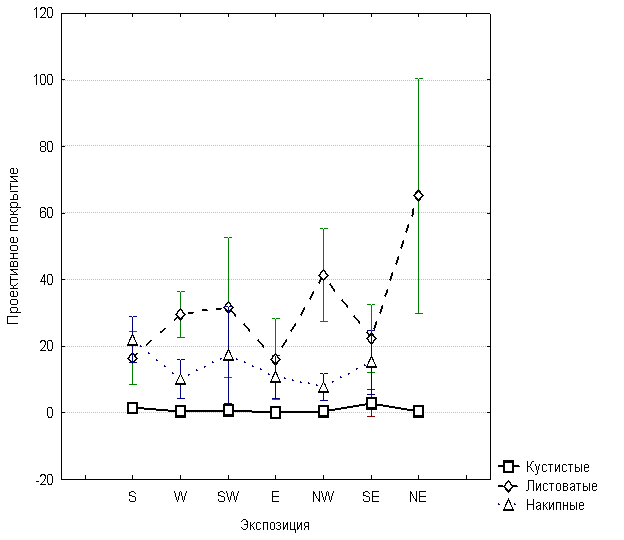
Рис. 3. Зависимость проективного покрытия разных жизненных форм лишайников от экспозиции ствола |
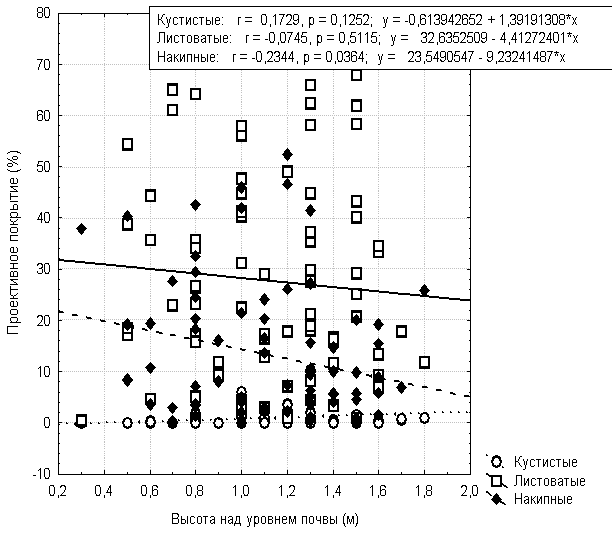
Рис. 4. Отношение проективного покрытия лишайников и высоты над уровнем почвы
|
В целом, на дубе зубчатом доминируют виды из семейства Physciaceae: Phaeophyscia hirtuosa, P. rubropulchra, Physconia detersa, P. subpulverulenta - в Хасанском районе, Phaeophyscia hispidula, P. hirtuosa, Physconia detersa - в Лазовском районе. Представители рода Heterodermia из того же семейства чаще встречаются также на этой породе.
На дубе монгольском доминируют виды из семейства Parmeliaceae: Myelochroa aurulenta, M. entotheiochroa - в Хасанском районе, - Flavoparmelia caperata, Myelochroa aurulenta, Parmotrema chinense - в Лазовском районе.
Помимо перечисленных видов, на обеих породах встречаются также другие доминанты, причем обнаруживаются существенные различия в их составе в зависимости от района исследования. На дубе зубчатом в Хасанском районе доминируют Caloplaca gordejevi, Lecanora allophana, Myelochroa entotheiochroa, а в Лазовском районе - Arthopyrenia stenospora, Biatora vernalis,Collema furfuraceum, Lecanora pachyheila. На дубе монгольском в Хасанском районе преимущественно встречаются Lecanora pachyheila, Lecidella elaeochroma, Schismatomma pericleum, Ramalina conduplicans, а в Лазовском - Buellia disciformis, Lecanora chlarotera, Ochrolechia parella. |
Таким образом, своеобразие в доминантных комплексах обнаруживается как в отношении каждого из двух видов дуба, так и в отношении каждого из двух исследованных районов. При этом, в разных районах исследования состав доминантов изменяется таким образом, что в Хасанском районе они одинаково часто встречаются на обеих породах, и это касается таких видов, как Caloplaca gordejevi, Lecanora allophana, Myelochroa entotheiochroa, Phaeophyscia rubropulchra. Напротив, в Лазовском районе общие виды на дубе монгольском и дубе зубчатом отсутствуют.
Чтобы выявить закономерности в изменении состава лишайниковых сообществ в разных районах более отчетливо, был проведен анализ попарного флористического сходства лихенометрических площадок c использованием метода информационно-флористического сходства, описанного в главе 3.
По результатам анализа информационно-флористического сходства площадок построены графы (рис. 5-9), на которых кружками с номерами обозначались лихенометрические площадки (вершины графа), а линиями разной толщины (ребра графа) - уровни сходства с пороговыми значениями 0.1 (10%) в интервале от 0.3 до 0.9. Площадки с уровнем сходства, равным единице, на графе показаны одним кружком. Сходные пробные площадки расположены ближе друг к другу. Чем больше общих видов у пары пробных площадок и чем выше их информативность, тем выше их сходство. Информативность редких видов в выборке описаний выше, чем информативность частых видов. Площадки объединены в группы при пороге сходства 0,5. Этот порог выбран методом ступенчатого анализа степени неравномерности графа по мере снижения порога сходства лих. площадок (Галанин, 1991; Беликович, 2001).
В тех случаях, когда индексов сходства, равных и более 0,5, в матрице было мало, при выделении групп, порог включения некоторых видов в группу понижался до 0,4. Все эти случаи далее оговариваются отдельно.
Первые три графа построены для эпифитных лишайников на дубе монгольском - два для Хасанского и один для Лазовского районов. Видовой состав на этой породе в Хасанском районе беднее, чем в Лазовском (66 и 89 видов соответственно).
Первый граф (рис. 5) мы рассматриваем как модель таксономической структуры населения эпифитных лишайников на дубе монгольском в Хасанском районе Приморского края (окр. п. Зарубино). Следует сказать, что таксономическая неоднородность населения лишайников, по всей вероятности, более устойчива во времени, чем неоднородность, выявленная с использованием проективного покрытия. На графе выделяется крупная группа площадок (40, 25, 49, 48, 75, 28, 3, 27, 4, 77, 33, 78, 67, 17,63, 39) со степенью связи 0,5 и выше. С такой же степенью связи выделились и другие группы. Эта группа лихенометрических площадок характеризуется следующей группой видов: Myelochroa entotheiochroa, M. aurulenta, Phaeophyscia rubropulchra, Ramalina conduplicans, R. dilacerata, Caloplaca gordejevi, Lecanora allophana, L. pachyheila, Lecidella elaeochroma, Schismatomma pericleum. Указанные виды являются доминирующими и обильными на дубе монгольском в месте исследования, их можно назвать «ядром» характерных видов. Здесь они имеют наибольшую частоту встречаемости и наибольшее проективное покрытие.
Кроме того, в эту группу входят Tephromela atra (отмечен на семи площадках) и Phaeophyscia hispidula (отмечен на шести площадках). Эти виды не являются доминирующими и обильными для дуба монгольского, но они также весьма характерны для него. Tephromela atra на дубе зубчатом нами вообще не найден, а Phaeophyscia hispidula чаще встречается на дубе монгольском, и при этом абсолютно доминирует на отдельных стволах его.
Остальные группы площадок характеризуются редко встречающимися в данной выборке видами или их комбинациями. Первая группа площадок (10, 59, 21, 45, 46, 57), помимо основных видов, характеризуется видами Myelochroa subaurulenta, Pyxine sorediata, Punctelia borreri. Эти виды не являются доминирующими и обильными, они встречаются периодически, причем являются более характерными для дуба монгольского, на котором имеют большее проективное покрытие.
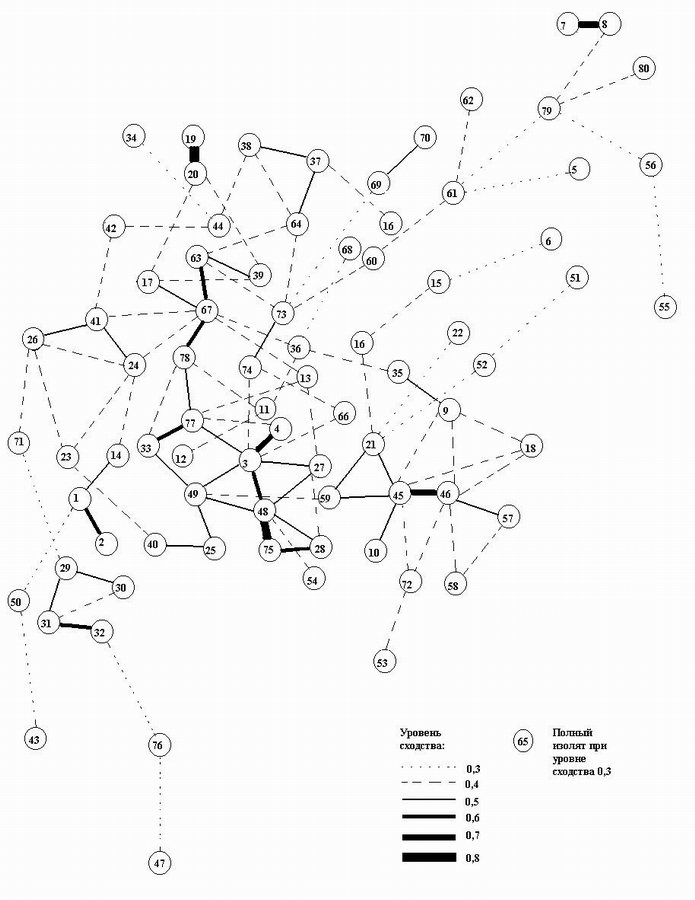
Рис. 5. Граф таксономической структуры населения эпифитных лишайников на дубе монгольском в Хасанском районе Приморского края. Обозначения даны в тексте.
Вторая группа лихенометрических площадок (29, 30, 31, 32) выделилась благодаря сочетанию видов Phaeophyscia hirtuosa, Ph. hispidula, Physconia detersa. Это своеобразная группа, так как Phaeophyscia hirtuosa и Physconia detersa доминируют с высоким проективным покрытием на дубе зубчатом, а на дубе монгольском в месте исследования являются редкими. Ph. hispidula, как отмечено выше, хотя и не является доминирующим видом на дубе монгольском, но встречается на нем гораздо чаще.
Третья группа площадок (1, 2, 14) выделилась за счет видов Heterodermia hypoleuca, H. diademata и H. speciosa. Первый из них чаще встречается на дубе монгольском, причем проективное покрытие его здесь выше, чем на дубе зубчатом. Второй вид, наоборот, чаще и с большим проективным покрытием встречается на дубе зубчатом. H. speciosa наблюдается одинаково нечасто на обоих видах дуба и не имеет высокого проективного покрытия.
Четвертая группа (26, 24, 41) характеризуется видами Pyxine sorediata, Anaptychia isidiata, Lecanora chlarotera и Parmotrema chinense. Все они намного чаще встречаются на дубе монгольском. Anaptychia isidiata отмечен на дубе зубчатом как достаточно редко встречающийся вид, на лихенометрических площадках не зарегистрирован. Pyxine sorediata в нашей выборке найден однажды.
Пятая группа (38, 37, 64) выделилась на основании характерных для дуба монгольского видов - Pertusaria submultipuncta, P.pertusa, Parmelia fertilis, Rinodina xanthophaea, Ochrolechia parella, Parmotrema chinense, Heterodermia speciosa. Это виды, за исключением Parmotrema chinense, с низким проективным покрытием. Они встречаются на обоих дубах нередко, за исключением Pertusaria pertusa и Ochrolechia parella, которые на дубе зубчатом в нашей выборке не отмечены. Rinodina xanthophaea на дубе зубчатом встречается гораздо реже, чем на дубе монгольском, хотя, возможно, это связано с тем, что на солнце в сухом состоянии этот вид слабо заметен на коре, и поэтому реже регистрируется. В дубняке же из дуба монгольского даже в солнечную погоду намного больше тени, и лишайники сохраняются более влажными.
Шестая группа (19, 20) характеризуется такими видами, как Flavoparmelia caperata, Tephromela atra, Ochrolechia parella. Седьмая группа (73, 74) выделилась видами Lecanora pulicaris, Punctelia borreri, Ochrolechia parella. Из этих видов Lecanora pulicaris наиболее редко встречается в нашей выборке и ни разу не отмечен на дубе зубчатом. Два других вида также более часто встречаются на дубе монгольском и имеют на нем большее проективное покрытие. Восьмая группа (69, 70) выделилась благодаря видам Buellia disciformis и Trapeliopsis viridescens. Первый вид - редкий в нашей выборке для дуба монгольского, а на дубе зубчатом он не обнаружен. Второй вид найден на обеих породах дуба, но также является редким. Последние три группы имеют одну особенность, состоящую в том, что выделившиеся пары площадок располагаются на одном стволе (каждая на своем), их видовой состав оказался более однородным по сравнению с другими стволами деревьев.
Девятая группа (9, 35) характеризуется видами Pyxine sorediata, Myelochroa subaurulenta и Parmelia fertilis. Особенности их распределения даны выше. Десятая группа (7, 8) выделилась благодаря видам Punctelia borreri, Heterodermia diademata, Anaptychia isidiata, Ochrolechia parella, Parmotrema stuppeum. В основном, эти виды больше характерны для дуба монгольского, за исключением Heterodermia diademata, который имеет большее проективное покрытие и встречаемость на дубе зубчатом. Parmotrema stuppeum является редким видом (найден однажды) в нашей выборке и только на дубе монгольском.
На графе есть один изолят (65) при уровне сходства 0,3, а также имеются изоляты при уровне сходства 0,4 - это площадки: 5, 22, 34, 43, 47, 50, 51, 52, 55, 56, 68 и 76, на графе их связь с остальными площадками показана при уровне сходства 0,3. В основном, это площадки с богатым видовым составом от 5 до 12 видов, только три из них имеют по 3 или 4 вида. Почти все площадки-изоляты выделились из-за наличия на них определенного, редкого в нашей выборке, вида лишайника, найденного только на этой площадке. Соответственно для площадок 5, 22, 43, 47, 51, 55, 68, 76, это такие виды как Heterodermia japonica, Lepraria sp., Lecanora argentata, Physconia distorta, Physconia sp., Pyxine sibirica, Ramalina roesleri, Rinodina exigua. Площадки 34 и 56 характеризуются наличием групп видов - это, соответственно, Heterodermia japonica, Physconia distorta, Pyxine sibirica и Heterodermia dissecta, Lecanora glabrata, Rinodina septentrionalis. Две площадки 50 и 52 имеют бедный видовой состав (4 и 5 видов соответственно), в который входят виды, характерные для дуба монгольского.
Второй граф (рис. 6) мы рассматриваем как модель таксономической структуры населения эпифитных лишайников на дубе монгольском в окрестностях пос. Кравцовка Хасанского района Приморского края. В состав древостоя здесь входит пихта черная (Abies nephrolepis).
Данная графовая модель построена при низком уровне сходства 0.3, так как в данной матрице мало индексов сходства выше 0,4 и 0,5. Отдельные кластеры выделены при уровне сходства 0.4 и включают мало описаний со степенью сходства более 0.5. Структура этого графа неопределенная, здесь нет главного кластера, который объединяет, как правило, большую группу площадок с ядром характерных видов для данного района исследований и данного субстрата. Выделившиеся кластеры с небольшим количеством площадок, объединены разными степенями сходства от 0.4 до 0.9.
Высоких степеней сходства между описаниями на графе наблюдается немного. Это говорит о том, что лишайниковый покров флористически слабо структурирован на уровне лишайниковых сообществ на дубе монгольском в исследованном районе. Вероятно, сказывается пограничное влияние и взаимодействие двух разных флор: сихотэ-алинской - с севера и корейской - с юга и, скорее всего, ядро характерных видов лишайников здесь состоит из видов лихенофлор разного генезиса, а лихеногруппы, вероятно, находятся в состоянии формирования. Здесь произрастают виды, которые не встречаются южнее, но появляются на севере Хасанского района и становятся обильнее севернее в составе Сихотэ-Алинской лихенобиоты. Это такие виды, как Caloplaca citrine, Ochrolechia yasudae, Myelochroa persidians, Pertusaria multipuncta, P. laevigata, P. leioplaca, P. subobducens, P. velata, Bacidia biatorina, Anzia stenophylla, Rinodina sophodes, Heterodermia microphylla, Phaeophyscia melanchra, P. primaria, P. squrrosa, Physconia grumosa. Виды Menegazzia terebrata, Graphis scripta, Phaeophyscia hispidula, Flavoparmelia caperata, Rinodina xanthophaea, Parmotrema chinense встречаются здесь с большей частотой и большим проективным покрытием, чем на юге Хасанского района; эти виды больше характерны для лесов из дуба монгольского.
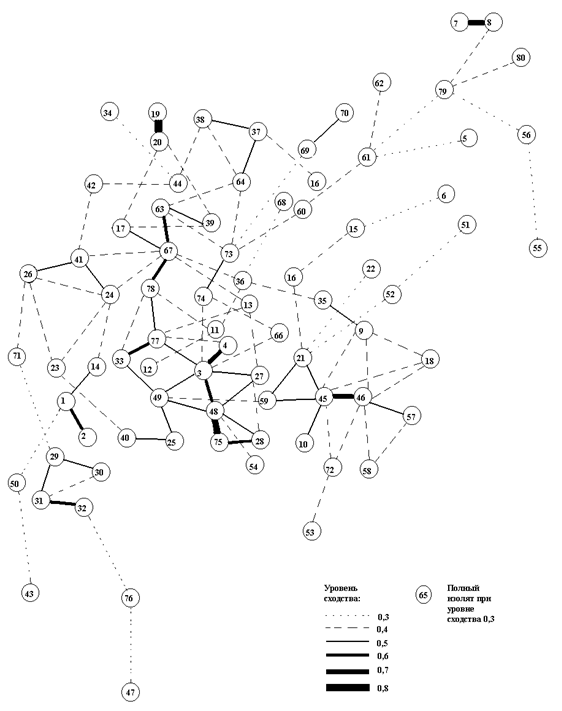
Рис. 6. Граф таксономической структуры населения эпифитных лишайников на дубе монгольском в окрестностях пос. Кравцовка Хасанского района Приморского края. Обозначения даны в тексте к первому графу.
Наибольший кластер на графе объединяет десять вершин (23, 24, 25, 69, 70, 29, 47, 30, 45, 33). Их характеризуют площадки с большим количеством видов (до 15). Объединены эти площадки за счет общих доминирующих видов для дуба монгольского в окрестностях пос. Кравцовка: Caloplaca gordejevi , Lecanora pachyheila , L . chlarotera , Myelochroa aurulenta , чуть реже (от 30% до 43% ) встречаются характерные виды Caloplaca citrina , Lecanora allophana , Myelochroa entotheiochroa , Parmotrema chinense , Phaeophyscia hispidula , Rinodina xanthophaea . Слабые связи между площадками, входящими в кластер, отражают группу видов с низкой встречаемостью и отсутствием корреляции друг с другом, что вносит неоднородность в кластер и граф в целом. Это связано с более высоким видовым разнообразием лихенофлоры в зоне контакта лихенофлор разного генезиса. К таким видам относятся Anzia stenophylla , Graphis scripta , Phaeophyscia squrrosa , P . sciastra , Pertusaria velata , Schismatomma pericleum ; они более характерны для дубняков из дуба монгольского. Из них на дубе зубчатом на юге Хасанского района и в Лазовском заповеднике отмечен только вид Schismatomma pericleum .
Вторая группа площадок (5, 6, 7, 21, 22) выделилась за счет видов Myelochroa entotheiochroa , Pertusaria subobducens , Lecanora pachyheila , Caloplaca gordejevi . Это - обычные для дуба монгольского и дуба зубчатого виды, за исключением вида Pertusaria subobducens , который не встречается на дубе зубчатом ни в Хасанском, ни в Лазовском районах. Этот вид нередко встречается в хвойно-широколиственных лесах (Чабаненко, 2002), на дубе монгольском в нашей выборке данный вид встречается именно в том месте, где дуб монгольский смешивается с пихтой черной, образуя смешанный лес. Вероятно, данная лихногруппа - одна из характерных для границы двух лихено-флористических провинций.
Следующая совокупность площадок (12, 15, 14, 17, 31) на графе отличается бедным видовым составом. Описания содержат по два, четыре вида, а именно Lecanora allophana , Phaeophyscia hispidula , Rinodina xanthophaea , Lepraria incana . Такое зарастание наблюдается в тех участках леса, где стволы очень высокие, старые, редко расположенные и степень сомкнутости полога леса максимальная. Виды Phaeophyscia hispidula , Rinodina xanthophaea и Lepraria incana зачастую выбирают подобные притененные участки ствола и в других изученных нами районах. Например, четвертая группа площадок (24, 25, 85) на дубе монгольском на юге Хасанского района тоже выделилась за счет присутствия на них Rinodina xanthophaea и Phaeophyscia hispidula . (см. первый граф на рис. 5).
Аналогичным путем выделилась группа вершин (1, 2, 3). Здесь также мало видов, но они характерны для дуба монгольского - Myelochroa aurulenta , Phaeophyscia hispidula , Lecanora pulicaris . Последний вид делает эту группу необычной, так как здесь он встречается редко и только на дубе монгольском в Хасанском районе, более обычен становится севернее.
Четвертая группа вершин (26, 27, 28) обособилась за счет редкого сочетания видов: Parmotrema stuppeum , Lepraria incana , Myelochroa entotheiochroa . Parmotrema stuppeum является редким, ранее он был найден однажды на дубе монгольском на юге Хасанского района.
На графе имеются две пары площадок (10 и 11) и (67 и 68) с большим сходством между ними и сильной изолированностью. Обе пары близко расположены и содержат редко встречающиеся здесь виды. Первая пара имеет низкое видовое разнообразие (5 видов) и отличается сочетанием видов Pertusaria submultipuncta , Myelochroa entotheiochroa , Lecanora allophana . Pertusaria submultipuncta чаще встречался на дубе зубчатом на юге Хасанского района, остальные два вида характерны для обоих видов дуба. Вторая пара, наоборот, имеет большое разнообразие видов (11). Эту пару отличает сочетание видов Heterodermia microphilla , H . speciosa , Ochrolechia pallescens , Parmelia saxatilis , Punctelia rudecta , Pyxine sorediata ; все они чаще встречаются на дубе монгольском, и здесь их проективное покрытие увеличивается.
Третий граф (рис. 7) мы рассматриваем как модель таксономической структуры населения эпифитных лишайников на дубе монгольском в окрестностях пос. Киевка Лазовского района Приморского края. Данная графовая модель построена при довольно низком уровне сходства 0.3. Отдельные кластеры выделены при уровне сходства 0.4 и включают мало пар площадок со сходством более 0.5. Это говорит о том, что флористическая структура населения лишайников эпифитов выражена не четко.
На графе выделяется группа из двадцати площадок (67, 57, 56, 49, 31, 63, 62, 22, 88, 43, 41, 82, 44, 85, 35, 90, 2, 74, 40, 36) с минимальным порогом сходства между ними 0.4. Эту группу площадок объединяет «ядро» характерных видов лишайников для дуба монгольского в Лазовском районе. К ним относятся наиболее распространенные здесь виды с высокой частотой встречаемости и высоким проективным покрытием - это доминанты Buellia disciformis , Caloplaca gordejevi , Flavoparmelia caperata , Lecanora chlarotera , L . allophana , Myelochroa aurulenta , Ochrolechia parella , Parmotrema chinense , Phaeophyscia rubropuchra , Ramalina dilacerata . Указанные виды больше характерны для дуба монгольского, чем для дуба зубчатого, предпочитают более притененные местообитания. На дубе зубчатом встречаемость и проективное покрытие некоторых из них гораздо ниже, а некоторые перестают встречаться вообще. Этот эффект усиливается на юге Хасанского района.
Общих доминирующих видов для обоих видов дуба в Лазовском районе нет. Слабое сходство между площадками (вершинами графа) в первом кластере объясняется большим количеством видов, которые сильно повышают общее флористическое разнообразие, но при этом их встречаемость в рассматриваемой выборке низкая. В данной группе это такие виды, как Cetrelia pseudolivetorum , Lecanora symmicta , L . subrugosa , Pertusaria leioplaca , Rinodina pyrina , R . sophodes , Heterodermia hypoleuca . Наблюдается существенная разница по количеству видов эпифитных лишайиков, встречающихся на обоих видах дуба на юге Хасанского района и в Лазовском районе, это соответственно 68 и 102 вида, зарегистрированных нами только на лихенометрических площадках.
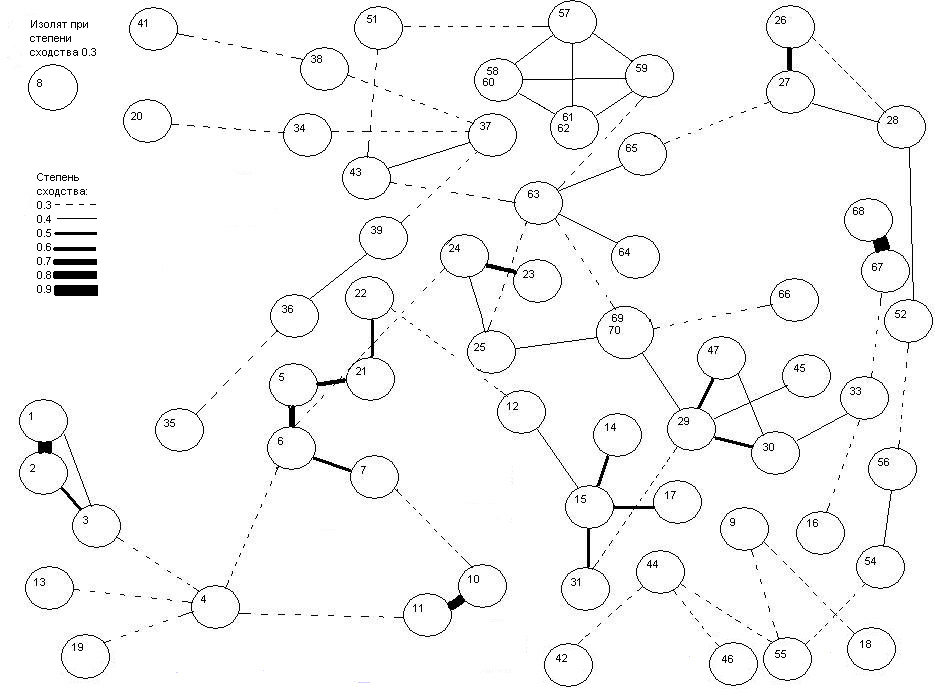
Рис. 7. Граф таксономической структуры населения эпифитных лишайников на дубе монгольском в окрестностях п. Киевка Лазовского заповедника Приморского края. Обозначения даны в тексте к первому графу.
Первый кластер несколько вытянут слева направо (связь между площадками 62 и 88), его можно разделить на два кластера, каждый из которых обособлен благодаря своей собственной группе видов лишайников. Первая подгруппа вершин (67, 57, 56, 49, 31, 63, 22, 62) в большинстве своем содержит накипные виды - Pertusaria multipuncta и P . pertusa . Вторая подгруппа вершин (88, 43, 41, 82, 44, 85, 35, 90, 2, 74, 40, 36) содержит листоватые виды - Parmelia fertilis и P . saxatilis .
Следующей рассмотрим группу вершин (16, 28, 9, 8, 10, 17, 11, 15, 14, 5, 21, 12), обособившихся во второй кластер, также при уровне сходства 0.4 между описаниями лихенометрических площадок. Эти описания содержат стабильный состав доминирующих видов лишайников для дуба монгольского в Лазовском районе. За счет них вершины имеют сильные связи, но они выделились на графе благодаря виду Menegazzia terebrata . Данный вид очень редко встречается на юге Хасанского района; он чаще начинает появляться в окрестностях пос. Кравцовка на севере Хасанского района, а в Лазовском районе уже выступает в роли элемента, формирующего особые лишайниковые сообщества, которые не образуются в Хасанском районе даже на дубе монгольском. Menegazzia terebrata произрастает в разных типах леса на разных видах деревьев и на камнях, но в дубняках предпочитает дуб монгольский и ближе к Сихотэ-Алиню заметно увеличивает свою активность.
Кроме двух больших кластеров, на графе выделились три группы и пять пар площадок. Уровень сходства здесь также от 0.4 и выше. Группы и пары площадок четко выделяются из основной массы описаний, имеющих уровень сходства 0.3. Группу вершин (19, 33, 27) объединяет комплекс накипных видов - Lecanora pulicaris , Pertusaria submultipuncta , Rinodina archaea ; все они в большей степени тяготеют к местообитаниям на дубе монгольском во всех исследованных районах.
Следующую группу вершин (61, 64, 65) объединяют накипные виды: Chrysothrix chlorina , Lecidella elaeochroma , Tephromela atra , они также в большей степени тяготеют к местообитаниям на дубе монгольском во всех исследованных районах.
Группу вершин (38, 37, 89) объединяет листоватый вид Parmelia saxatilis . Этот вид в Лазовском районе встречается чаще - увеличивает свою встречаемость от 1% на юге Хасанского района до 21% в Лазовском районе. P . saxatilis чаще встречается на дубе монгольском, чем на дубе зубчатом во всех районах исследования. В остальном же описания площадок имеют несколько разрозненный видовой состав, и выделить конкретную лихеногруппу для них не представляется возможным.
Рассмотрим пары площадок (81 и 78, 3 и 84, 30 и 23, 25 и 66), выделившиеся при уровне сходства более 0.4. Все эти пары не являются близкорасположенными. Первая пара площадок на графе обособилась за счет видов Caloplaca cerina , Phaeophyscia stellaris , Tephromela atra , вторая - за счет видов Usnea diplotypus , Ramalina roesleri , третья - за счет видов Lecidella elaeochroma , Pertusaria multipuncta , Pyxine sorediata , Rinodina pyrina , четвертая - за счет видов Chrysothrix chlorina , Lepraria incana , Phaeophyscia hispidula .
На графе имеются изоляты при уровне сходства 0,3 - это площадки 1, 13, 75, 76, и группа площадок 52, 53, 54. Каждая из них выделилась из-за наличия определенного вида лишайника, редкого в данной выборке. На площадке 1 найден вид Parmelia adaugescens , который, возможно, не является редким на дубе монгольском в Лазовском районе, так как для Сихотэ-Алиня его приводят как частый (Скирина, 1995), но в нашей выборке данный вид незначительно представлен в Лазовском районе только на дубе монгольском. На дубе зубчатом в нашей выборке Parmelia adaugescens здесь не отмечен совсем. На площадке 13 найден вид Evernia mesomorpha , который не является редким на дубе монгольском несколько севернее в Лазовском районе, но в приморских южных дубняках он встречен нами единично и только на дубе монгольском, в Хасанском районе не найден. На площадке 75 найден новый вид из рода Caloplaca sp ., возможно новый для Приморского края. На площадке 76 найден также вид Evernia mesomorpha и редкий вид Usnea rubicunda .
Группа площадок (52, 53, 54) оказалась изолирована (уровень сходства с остальными площадками менее 0,2). Она имеет очень бедный видовой состав, здесь встречается всего три вида - Myelochroa entotheiochroa , M . aurulenta , Phaeophyscia hispidula , все они более характерны для дуба монгольского, чем для дуба зубчатого.
В целом, можно сказать, что видовой состав и ядра характерных видов и отдельных лихеногрупп включают виды, характерные для дуба монгольского. Нет ни одной лихеногруппы, состоящей из видов более характерных для дуба зубчатого. Таким образом, в Лазовском районе мы наблюдаем иную картину состава и распределения ядра характерных видов и лихеногрупп на двух видах дуба, в сравнении с более южными и западными дубняками в Хасанском районе. В Хасанском районе наблюдается большее проникновение видов, характерных для дуба зубчатого, в состав лихеногрупп на дубе монгольском.
Кроме разницы в количественном составе видов, «ядро» характерных для дуба монгольского видов лишайников существенно меняется в Лазовском районе. Общими для «ядер» лишайниковых сообществ в обоих районах являются только Caloplaca gordejevi , Lecanora allophana , Myelochroa aurulenta и Phaeophyscia rubropulchra , тогда как здесь наиболее распространены с высокой частотой встречаемости и высоким проективным покрытием Buellia disciformis , Flavoparmelia caperata , Lecanora chlarotera , Ochrolechia parella , Parmotrema chinense , Ramalina dilacerat а .
Третий граф, связанный с эпифитными лишайниками на дубе монгольском в окр. п. Кравцовка на севере Хасанского района, демонстрирует отсутствие главного кластера, который объединяет, как правило, большую группу площадок с ядром характерных видов. Кроме того, здесь появляются виды, более характерные для «сихотэ-алинской» лихенофлоры.
Сравнение лишайниковых сообществ на дубе монгольском в трех районах позволяет рассматривать две обособившиеся лихенобиоты с характерными ядрами видов на самом юге Хасанского района и в Сихотэ-Алинских горах Лазовского района. Третье местообитание находится в области широкого распространения дубовых и широколиственных лесов и характеризуется достаточно разнообразным видовым составом, но низкой связанностью видов лишайников на уровне сообществ.
Следующие два графа построены для эпифитных лишайников на дубе зубчатом - в Хасанском и Лазовском районах. Видовой состав на этой породе, как и в случае дуба монгольского, беднее в Хасанском районе, чем в Лазовском (52 и 67 видов соответственно).
Четвертый граф (рис. 8) рассматривается как модель таксономической структуры населения эпифитных лишайников на дубе зубчатом в Хасанском районе Приморского края. На графе ясно выделяется крупная группа площадок (52, 21, 23, 53, 18, 30, 33, 10, 40, 41, 62, 61, 77, 31, 37, 13, 45, 39, 34, 32, 6, 87, 47, 59, 19, 86, 63, 48, 54, 83, 79, 78) с минимальной степенью сходства между ними (0,5). Эту группу площадок объединяет «ядро» характерных видов лишайников для дуба зубчатого. К ним относятся наиболее распространенные здесь виды с высокой частотой встречаемости и высоким проективным покрытием: Phaeophyscia hirtuosa , P . rubropulchra , Physconia detersa , Myelochroa entotheiochroa , Caloplaca gordejevi , Lecanora allophana. На дубе монгольском из этих видов доминируют Myelochroa entotheiochroa , Caloplaca gordejevi , Phaeophyscia rubropulchra ; эти виды являются общими доминирующими видами для обоих видов дуба.
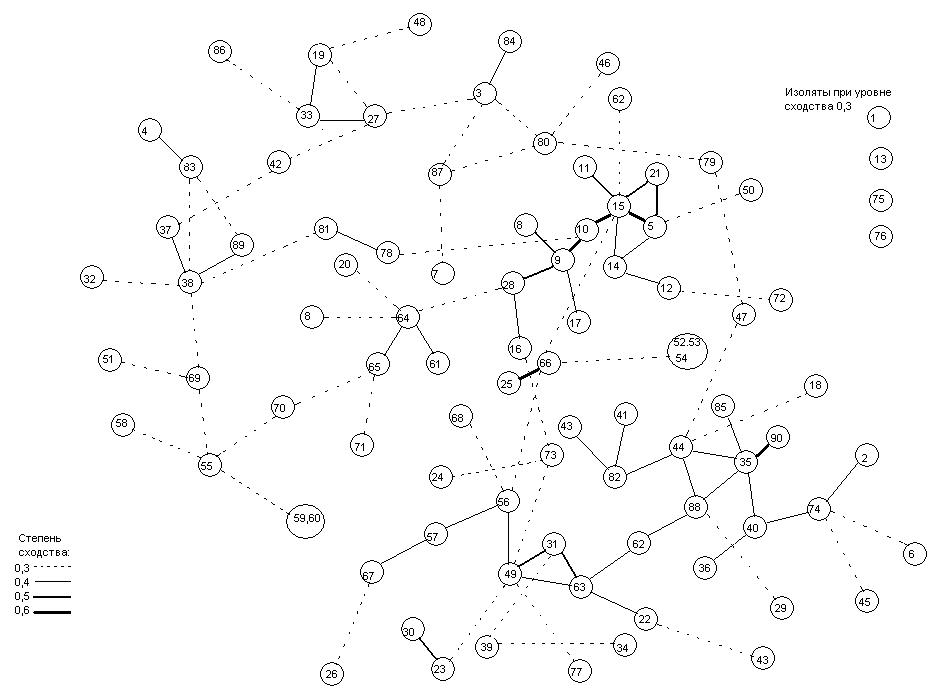
Рис. 8. Граф таксономической структуры населения эпифитных лишайников на дубе зубчатом в Хасанском районе Приморского края.
Обозначения даны в тексте к первому графу.
Кроме ядра видов, на графе выделились группы различной величины. Первая самая большая группа включает девять площадок (57, 70, 36, 29, 2, 72, 71, 38, 74) с минимальной связью 0.5 и максимальной 0.7. Данная группа выделилась из-за пары видов Physconia kurokawae и P . subpulverulenta . В нашей выборке первый вид на дубе монгольском отсутствует, а второй найден дважды несмотря на то, что на дубе зубчатом они часто встречаются вместе.
Вторая группа включает всего четыре площадки (80, 81, 60, 12), но она четко характеризуется наличием редких на дубе зубчатом видов: Lecanora pachyheila , Pyxine sibirica , Melanelia huei и Phaeophyscia hispidula . Первый вид часто и обильно встречается на дубе монгольском. Pyxine sibirica чаще отмечена на дубе монгольском, хотя в целом встречается не часто и не обильно. Последние два вида на дубе монгольском не частые, но Phaeophyscia hispidula на отдельных стволах имеет очень высокое проективное покрытие, чего нет на дубе зубчатом.
Третья группа площадок (82, 84, 8, 9) выделилась на основе очень слабого зарастания. Это площадки, отражающие состояние лишайников на тех участках ствола, которые регулярно обгорают. На таких участках главным образом встречаются Phaeophyscia hirtuosa , P . rubropulchra , Physconia detersa . Эти площадки находятся на разной высоте ( 60 см , 1 м , 1,2 м , 1,4 м ), а также имеют разную экспозицию (западную, восточную, северо-восточную).
Четвертая группа площадок (24, 25, 85) выделилась за счет присутствия на них Rinodina xanthophaea и Phaeophyscia hispidula . Первый вид в нашей выборке встречается на дубе зубчатом реже (6 раз), чем на дубе монгольском (16 раз). Особенности распределения второго вида приведены выше.
Пятая группа (пара площадок 26 и 27) выделилась благодаря наличию на них видов Bacidia circumspecta и Lecanora pachyheila . Эти площадки расположены на одном стволе, близко друг к другу и могут считаться за одну. Можно рассматривать эту пару как изолят с редким видом Bacidia circumspecta . Однако данный вид довольно мелкий, накипный, поэтому трудный для визуальной регистрации, и, возможно, встречается чаще, чем он отмечен в нашей выборке. Второй вид часто и обильно встречается на дубе монгольском
Кроме того, интересна шестая группа, состоящая из двух площадок (49 и 50) с богатым видовым составом (12 и 15 видов). Здесь встречаются редкие и малообильные в нашей выборке для дуба зубчатого виды: Lecanora chlarotera , L . pachyheila , Rinodina xanthophaea , Candelaria concolor и Pertusaria submultipuncta . Все эти виды чаще встречаются на дубе монгольском. В целом, виды из родов Pertusaria , Lecanora , Ochrolechia намного обильнее на дубе монгольском. Candelaria concolor встречается не часто, но на отдельных стволах имеет высокое обилие. Кроме того, данная группа характеризуется видом Physconia kurokawae , который характерен для дуба зубчатого и имеет здесь большее проективное покрытие и встречаемость.
На графе имеются также изоляты при уровне сходства 0.4 - это площадки 4, 16, 42 и 51. Каждая из них выделилась из-за наличия определенного вида лишайника, встреченного только на этой площадке. На площадке 4 найден вид Parmelia saxatilis , здесь довольно редкий, а на дубе монгольском в нашей выборке он не отмечен совсем. Площадка 16 характеризуется наличием вида Pyxine sorediata , который представляет собой один из редко встречающихся здесь видов, хотя на дубе монгольском он был найден в большем количестве. Оба вышеуказанных вида отмечаются гораздо чаще севернее, в Сихотэ-Алине, на хвойных и широколиственных породах деревьев, а также на замшелых камнях (Скирина, 1995).
Площадка 42 характеризуется наличием вида Physconia distorta . Он редко встречается на дубе зубчатом, и на дубе монгольском, хотя для Сихотэ-Алиня приводится как часто встречающийся (Скирина, 1995). Площадка 51 характеризуется наличием вида Lecanora argentata , который редко встречается как на дубе зубчатом, так и на дубе монгольском. Для Сихотэ-Алиня этот вид приводится также как редко встречающийся (Скирина, 1995).
Следует отметить, что группы площадок, выделившиеся на графе для дуба зубчатого, помимо главной группы с «ядром» характерных видов, характеризуются видами, свойственными выборке с дуба монгольского, то есть видами, имеющими на нем большее проективное покрытие и большую встречаемость. В то же время из групп площадок, выделившихся на графе для дуба монгольского, только одна группа характеризуется видами, свойственными выборке с дуба зубчатого.
Пятый граф (рис. 9) рассматривается как модель таксономической структуры населения эпифитных лишайников на дубе зубчатом в Лазовском заповеднике Приморского края. Как видно из рисунка, данная графовая модель имеет довольно слабо выраженную структуру при уровне сходства между площадками более 0.4. Здесь мало описаний объединено в кластеры с высокой степенью сходства.
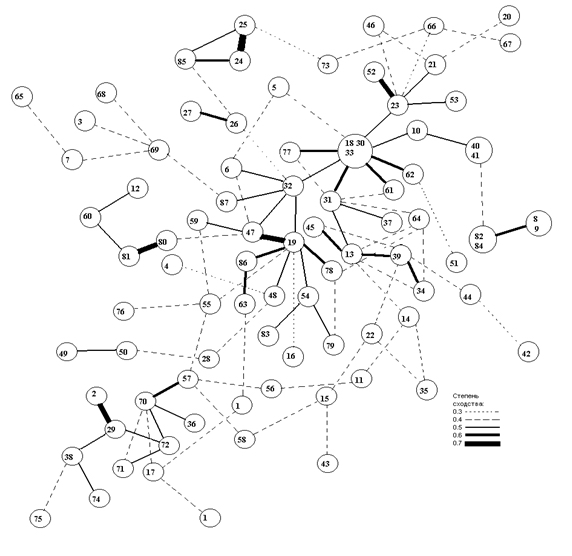
Рис. 9. Граф таксономической структуры населения эпифитных лишайников на дубе зубчатом в Лазовском заповеднике Приморского края.
Обозначения даны в тексте к первому графу.
На графе выделяется группа из 16 площадок (3, 17, 18, 73, 74, 10, 11, 34, 55, 32, 13, 48, 56, 57, 36, 19) с минимальной степенью сходства между ними 0.5. Эту группу площадок и массу связанных с ними площадок со степенью сходства 0.4 объединяет «ядро» характерных видов лишайников для дуба зубчатого в Лазовском заповеднике. К ним относятся наиболее распространенные здесь виды с высокой частотой встречаемости и высоким проективным покрытием: Arthopyrenia stenospora, Biatora vernalis , Collema furfuraceum , Lecanora pachyheila , Phaeophyscia hispidula , Ph . hirtuosa , Physconia detersa . На дубе монгольском в Лазовском районе ни один из этих видов не доминирует и встречаются они там изредка (например, Biatora vernalis , Collema furfuraceum ), либо не встречены вообще. Однако Phaeophyscia hirtuosa и Physconia detersa также входят в состав ядра характерных для дуба зубчатого видов в Хасанском районе. Таким образом, здесь сохраняется, независимо от района, общая доминантная группа - основа населения лишайников на дубе зубчатом. Сюда относятся листоватые лишайники из семейства Physciaceae , они имеют большее проективное покрытие, чем другие виды-доминанты, которые являются в основном накипными. Исключение представляет Collema furfuraceum , но данный вид никогда не имеет большого проективного покрытия.
Кроме ядра видов, на графе выделились группы площадок различной величины. Первая группа включает четыре площадки (53, 12, 9, 7) с минимальной связью 0.5, и выделена благодаря группе видов - Caloplaca citrina, Myelochroa entotheiochroa , Phaeophyscia squarrosa , Physconia kurokawae .
В нашей выборке эти виды чаще встречались на дубе монгольском, но, тем не менее, вид Physconia kurokawae - более характерен для дуба зубчатого. Виды Caloplaca citrina и Phaeophyscia squarrosa обильно встречаются в окрестности п. Кравцовка Хасанского района, на юге же Хасанского района в больших массивах дуба зубчатого эти виды редки.
Вторая группа включает площадки (68, 65, 23, 30), она характеризуется сочетанием видов, чаще и обильно встречающихся на дубе монгольском: Caloplaca gordejevi, Melanelia huei , Myelochroa aurulenta , M. entotheiochroa, Rinodina septentrionalis, R. xanthophaea .
Третья группа площадок (44, 45, 50) выделилась за счет главным образом накипных видов - Caloplaca gordejevi, C. citrina, Lecanora pulicaris, Lepraria incana , которых на дубе зубчатом всегда мало и они не обильны, а на дубе монгольском - это обычные виды, часто обильные с большим проективным покрытием.
На графе (рис. 9) есть две пары площадок, которые имеют высокое сходство друг с другом. Они также выделились за счет видов, не характерных для дуба зубчатого. Пара площадок (6, 49) отличается наличием видов Heterodermia speciosa , Flavoparmelia caperata , Melanelia huei. Другая пара (41, 42) определяется видами Physconia subpulverulenta , Pyxine sorediata , Ramalina conduplicans . Эти площадки расположены на одном стволе, близко друг к другу; их можно рассматривать как изолят с редкими на дубе зубчатом в этом районе видами Pyxine sorediata , Ramalina conduplicans и вполне характерным для дуба зубчатого видом Physconia subpulverulenta .
Из приведенных характеристик лихеносинузий видно, что состав их сформирован главным образом видами, наиболее распространенными и обильными на дубе монгольском с примесью видов лишайников, характерных для дуба зубчатого. Данный лес из дуба зубчатого в Лазовском заповеднике находится в окружении огромного массива из дуба монгольского, который, в свою очередь, и оказывает такое мощное влияние на ценотическое формирование в покрове лишайников на дубе зубчатом. Возможно, виды, постоянно привносимые из массива дуба монгольского, изменяют ход развития лихеносинузий на дубе зубчатом в Лазовском районе.
Такое смешение видов отражается на графовой модели в виде малого количества площадок с высоким сходством и большим количеством площадок с малым сходством между описаниями. Поэтому на графе видно довольно однородное распределение описаний без четкого обособления лихенометрических описаний в кластеры. Здесь намечаются некие намеки на таксономическую структуру.
При сравнении лихеногруппировок на дубе зубчатом, как и на дубе монгольском выделяются «ядра» характерных видов. Среди них общими доминантами являются Phaeophyscia hirtuosa и Physconia detersa , в Хасанском районе также доминируют Caloplaca gordejevi , Lecanora allophana , Myelochroa entotheiochroa , Phaeophyscia rubropulchra , в Лазовском - Arthopyrenia stenospora , Biatora vernalis , Collema furfuraceum , Lecanora pachyheila , Phaeophyscia hispidula .
Если сравнить «ядра» видов лишайников на обоих дубах в каждом из районов, то в Лазовском районе нет сходства, а в Хасанском одинаково доминируют на дубе монгольском и дубе зубчатом Caloplaca gordejevi , Lecanora allophana , Myelochroa entotheiochroa , Phaeophyscia rubropulchra . Это говорит о большей давности ассоциаций обоих видов дуба на самом юге Приморского края и о большей обособленности лишайниковых сообществ данного района: они входят в состав «корейской» лихенобиоты, отличающейся от «сихотэ-алинской».
Группы видов лишайников-эпифитов, выделенные с помощью графовых моделей (рис. 5 и 6), можно проанализировать на предмет их связи с определенными параметрами, характеризующими местообитание. Эти данные представлены в таблице 4. Для каждого характерного вида приводится класс встречаемости в группе: I - 10-20 %, II - 20-40%, III - 40-60%, IV - 60-80%, V - 80-100%. Приведены данные по экспозиции площадок, их высоте на стволе и их количестве в группе.
Таблица 4.
Группы лихенометрических площадок, выделившиеся на графе 1 и 2, и виды лишайников с классом встречаемости
в этих группах 1
Punctelia borreri |
. |
IV |
. |
. |
. |
. |
. |
V |
. |
. |
V |
. |
. |
. |
. |
. |
. |
. |
Pyxine sorediata |
. |
IV |
. |
. |
V |
. |
. |
. |
. |
V |
. |
. |
. |
. |
. |
. |
. |
. |
Ramalina dilacera ta |
V |
IV |
. |
. |
V |
II |
. |
. |
III |
. |
III |
. |
. |
. |
. |
. |
. |
. |
Phaeophyscia hispidula |
III |
III |
V |
. |
. |
. |
. |
. |
. |
. |
. |
I |
. |
IV |
II |
IV |
. |
. |
Phaeophyscia hirtuosa |
. |
. |
V |
. |
. |
. |
. |
. |
. |
. |
. |
V |
V |
V |
III |
V |
III |
V |
Physconia detersa |
. |
. |
IV |
. |
. |
. |
. |
. |
. |
. |
. |
IV |
. |
IV |
. |
IV |
III |
. |
Heterodermia hypoleuca |
. |
. |
III |
V |
II |
II |
. |
. |
. |
. |
. |
I |
II |
. |
. |
. |
. |
. |
Heterodermia diademata |
. |
. |
. |
IV |
. |
. |
. |
. |
. |
. |
V |
I |
. |
II |
. |
. |
III |
. |
Heterodermia speciosa |
I |
. |
. |
IV |
. |
IV |
. |
III |
III |
. |
. |
. |
. |
. |
. |
. |
. |
. |
Lecanora chlarotera |
I |
II |
II |
II |
IV |
II |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
V |
Группа лих. площадок на графе |
Ядро видов |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
Ядро видов |
1 |
2 |
3 |
4 |
5 |
6 |
Экспозиция лих. площадки |
w,s,e,
n-w,
s-e |
s,
n-e,
w,e |
e |
s-e |
s,s-w,w |
s-e,s |
w |
s-w |
w |
s,s-e |
s-e |
e,w,
n-e,
n,s,s-e |
n,
n-e,
w,e,
s-e,
n-w |
n,n-e |
n-e,
w,e |
n-e,e |
n-e |
n,n-e |
Субстрат / вид дерева |
Qm |
Qm |
Qm |
Qm |
Qm |
Qm |
Qm |
Qm |
Qm |
Qm |
Qm |
Qd |
Qd |
Qd |
Qd |
Qd |
Qd |
Qd |
Средняя высота / м |
1,3 |
1,2 |
1,4 |
1,4 |
1 |
1,1 |
1,1 |
0,4 |
1,1 |
1,2 |
1,2 |
1,1 |
1,1 |
1,4 |
1,1 |
1,3 |
1,4 |
1,5 |
Число лих. площадок |
16 |
6 |
4 |
3 |
3 |
3 |
2 |
2 |
2 |
2 |
2 |
32 |
9 |
4 |
4 |
3 |
2 |
2 |
Виды |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Parmotrema chinense |
I |
III |
. |
. |
V |
IV |
. |
. |
. |
III |
III |
I |
II |
. |
. |
. |
. |
. |
Schismatomma pericleum |
III |
. |
. |
. |
V |
II |
. |
. |
V |
. |
. |
. |
. |
. |
. |
. |
. |
. |
Anaptychia isidiata |
. |
. |
. |
II |
IV |
. |
. |
. |
. |
. |
V |
. |
. |
. |
. |
. |
. |
. |
Ochrolechia parella |
I |
. |
II |
. |
II |
V |
V |
V |
. |
III |
V |
. |
. |
. |
. |
. |
. |
. |
Parmelia fertilis |
I |
. |
. |
. |
. |
V |
. |
. |
. |
V |
. |
I |
. |
. |
. |
. |
. |
III |
Pertusaria submultipuncta |
. |
. |
. |
. |
II |
V |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
V |
Pertusaria pertusa |
I |
. |
. |
. |
. |
IV |
. |
III |
III |
. |
. |
. |
. |
. |
. |
. |
. |
. |
Rinodina xant h ophaea |
II |
. |
II |
. |
II |
V |
. |
III |
. |
. |
. |
I |
. |
. |
. |
V |
. |
V |
Tephromela atra |
III |
. |
. |
. |
. |
. |
V |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
Flavoparmelia caperata |
. |
. |
. |
. |
. |
. |
V |
. |
. |
III |
III |
. |
. |
. |
. |
. |
. |
. |
Ochrolechia pallescens |
. |
. |
. |
II |
II |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
Lecanora pulicaris |
I |
. |
. |
II |
. |
II |
. |
V |
III |
. |
. |
. |
. |
. |
. |
. |
. |
. |
Buellia disciformis |
. |
. |
. |
. |
. |
. |
. |
. |
V |
. |
. |
. |
. |
. |
. |
. |
. |
. |
Trapeliopsis viridescens |
. |
. |
. |
. |
. |
. |
. |
. |
V |
. |
. |
I |
. |
. |
. |
. |
. |
. |
Parmotrema stuppeum |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
V |
. |
. |
. |
. |
. |
. |
. |
Pyxine sibirica |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
III |
I |
I |
III |
. |
. |
. |
. |
Leptogium burnetiae |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
II |
. |
. |
. |
. |
Physconia subpulverulenta |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
II |
III |
II |
. |
II |
. |
. |
Physconia kurokawae |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
I |
IV |
II |
. |
. |
III |
V |
Melanelia huei |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
I |
. |
V |
. |
. |
. |
. |
Candelaria concolor |
. |
. |
III |
. |
. |
. |
. |
. |
. |
. |
. |
II |
II |
. |
. |
. |
. |
V |
Collema subnigrescens |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
I |
. |
. |
. |
. |
. |
III |
Heterodermia japonica |
. |
I |
II |
. |
. |
II |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
III |
Rinodina exigua |
. |
II |
II |
. |
. |
. |
. |
. |
. |
. |
. |
I |
. |
. |
. |
. |
. |
. |
Lecanora glabrata |
I |
. |
. |
. |
. |
II |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
Lecanora sp. |
. |
. |
. |
. |
. |
II |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
Lepraria incana |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
I |
. |
. |
. |
. |
. |
. |
Rinodina archea |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
I |
. |
. |
. |
. |
. |
. |
Caloplaca cerina |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
I |
. |
. |
. |
. |
. |
. |
Rinodina pyrina |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
I |
I |
. |
. |
. |
. |
. |
Graphis scripta |
. |
I |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
Молодые талломы |
II |
I |
III |
II |
. |
IV |
. |
III |
III |
III |
III |
. |
. |
. |
. |
. |
. |
. |
Отмершие талломы |
IV |
III |
III |
II |
IV |
II |
. |
III |
V |
V |
V |
IV |
V |
III |
III |
. |
V |
V |
Обозначени в таблицах. Виды деревьев (субстрат): Qm - дуб монгольский, Qd - дуб зубчатый. Класс встречаемости площадок в группе: I - 10-20%, II - 20-40%, III - 40-60%, IV - 60-80%, V - 80-100%.
В таблице также приведена группа видов, которые имеют низкую встречаемость во всех выделенных группах. Это такие виды , как Ochrolechia pallescens, Heterodermia japonica, Rinodina exigua, R. archea, R. pyrina, Lecanora glabrata, Lecanor a sp., Lepraria incana, Caloplaca cerina, Graphis scripta.
Виды Cetrelia braunsiana, C. pseudolivetorum, Heterodermia dissecta, Heterodermia sp., Lecanora septentrionalis, Parmelia laevior, P. lobulifera, Physconia sp., Ramalina roesleri, Rinodina septentrionalis, Lepraria sp. в таблице не приведены , они оказались редкими в данной выборке . Вышеназванные виды отмечены на лихенометрических площадках, не вошедших ни в одну из выделившихся групп на графах 1 и 2, так как эти площадки имеют низкую степень сходства со всеми остальными площадками.
Проведенное исследование показало применимость метода лихенометрических площадок для анализа эколого-ценотической структуры населения эпифитных лишайников. Размер площадок должен меняться в зависимости от целей исследования: для выявления ценотических эффектов площадки должны быть меньшего размера (25-225 кв. см), а для выявления межвидовых корреляций, обусловленных эдафически, площадки должны быть большего размера (500-1000 кв. см). Оптимальный размер лихенометрической площадки должен зависеть и от размеров особей лишайников.