Вы находитесь: НАША БОТАНИЧКА  РАСТИТЕЛЬНОСТЬ МИРА Растительность Даурии Растительность Даурии как динамическая система
РАСТИТЕЛЬНОСТЬ МИРА Растительность Даурии Растительность Даурии как динамическая система
© Беликович А.В. Растительность Даурии [Электронный ресурс] – Владивосток, 2017. – http://ukhtoma.ru/geobotany/dahuria_11.htm
Введение || Флора Даурии || Провинции Даурии || Горные тундры, сообщества гольцового и подгольцового поясов || Леса: Кедровые, еловые, пихтовые леса || Сосновые леса || Лиственничные леса || Березовые и осиновые леса || Тополевые, ивовые, ольховые леса || Ильмовые рощи, ильмовые саванны, ильмовые харганаты || Кустарниковые сообщества: Абрикосовые, спирейные, миндальные харганаты и караганники || Ерники и ивняки || Степи: Типичные степи || Петрофитные степи и группировки литосистем || Даурские прерии || Сазовые степи и другие галофитные сообщества || Луга: Важные, сухие, остепненные луга || Сырые, болотистые луга. Болота || Пойменные комплексы || Растительность Даурии как динамическая система || Использованная литература
|
Растительность Даурии как динамическая система
Даурия как ботанико-географическая подобласть является громадным буфером между Восточносибирской мерзлотной тайгой, Центральноазиатскими пустынями, Южно-Сибирскими горными системами с Байкалом и Дальневосточными лесами муссонного типа. Поэтому здесь мы наблюдаем смешение типов растительности, характерных для разных широтных и меридиональных зон, и неуловимое равновесие, регулярно сдвигающееся в ту или иную стоорну в зависмости от преобладающего фактора климатоформирования. Такое уникальное соединение осложняется наличием крупных горных массивов (прежде всего возвышающейся в центре Даурии горной системы Хэнтея) и многочисленных невысоких хребтов почти широтного простирания. В этом смысле Даурия очень близка с двумя своими соседями – восточной и западной: Прибайкальем и Манчжурией, которые находятся в сходных условиях "контакта аридного и гумидного поясов, атлантического и пацифического океанических влияний, а также взаимодействий бореального холодного и центрально-азиатского континентального и сухого воздушных фронтов" (Намзалов, 2012).
Даурия является центром географического перекрестка, с севера которого расположена Якутия, с востока – Приамурье, с юга – пустыня Гоби, с запада – Байкал и окружающие его горные системы. По этому перекрестку в эпохи смены глобального климата путешествовали виды в направлении север – юг и запад – восток. В эпохи усиления азиатского антициклона область высокого давления над Даурией выдавливала влажные атмосферные массы на периферию, в связи с чем по восточной окраине Даурии в бассейне р. Хайлар (Аргунь) формировался регион с гумидным климатом и наличием многочисленных видов дальневосточного облика. В эпохи нарастания океаничности усиливалась циклональная деятельность тихоокеанского муссона, Даурию заселяли дальневосточные виды. Таким же образом колебалась граница между зонами влияния североазиатского и центральноазиатского антициклонов: в эпохи давления северных холодных атмосферных масс шла миграция на юг якутских и восточносибирских видов. Большое количество ксерофильных видов путешествовало по горным хребтам, имеющим меридиональное направление.
Именно в такой обстановке неравновесности в Даурии сформировался уникальный комплекс лесостепи, вовлекший в сферу своего влияния все территориально соседние таежные, степные и пустынные комплексы растительности. Именно лесостепь как феномен стоит в центре всех проблем изучения растительности Даурии, определяя не только современный облик этой территории, но и основные векторы возможного изменения ландшафта в будущем. Именно анализ лесостепи дает ключ для понимания процессов в растительности всей Даурии.
Проблемы образования растительности лесостепей и формирования их флоры неоднократно поднимались и разбирались как для Сибири (Лапшина, 1963; Михайлов, Мартынова, 1970; Намзалов, 1996; Банникова, 1998, 2003; Коротков, Краснощеков, 1998; Соколова, 2003; Басхаева, 2003; Намзалов, Басхаева, 2006; Антипова, 2006; Бальжинова, 2011; Макунина, 2014; Банаева, Намзалов, 2015; Цыренова и др., 2015, и др.) и Монголии (Горная лесостепь, 1983), так в частности и для Даурии (Рещиков, 1973, 1975; Дулепова, 1993, 2004; Корсунов и др., 2002; Пешкова, 2010; Намзалов и др., 2012; Холбоева и др., 2013; Королюк и др., 2013, и др.).
Согласно принятой в настоящее время в геоботанике концепции (Намзалов и др., 2012; Макунина, 2014; и др.), горная лесостепь считается высотной ступенью, растительность которой представляет сочетание травяных лесов на теневых склонах и степей и остепненных лугов – на южных склонах. Вслед за З.В. Карамышевой (1993), мы называем ее экспозиционной степью. Фактически, если рассматривать лесостепь как территориальное подразделение, это есть мезогеохора (Сочава, 1978), растительность которой имеет ранг мезокомбинаций (Грибова, Исаченко, 1972). Лесостепные мезокомбинации образуют высотный пояс в горах северного обрамления Центральной Азии (Намзалов, 1996; Намзалов, Басхаева, 2002).
Специфика лесостепи как биома заключается в ее
современном семиаридном характере, а в прошлом в постоянном балансировании на лезвии аридность – гумидность. В этом регионе, климат которого зависит от взаимодействия атмосферных процессов, идущих со стороны Тихого, Атлантического, Северного Ледовитого океанов и пустынь Центральной Азии, наблюдается крайне неустойчивое равновесие, готовое сдвинуться в любую сторону в зависимости от преобладания центра влияния. Здесь мы имеем тесный контакт лесной, степной и луговой растительности, который способствовал созданию пограничных типов – даурских прерий и даурского харганата.
Для понимания природы лесостепи следует вкратце пробежаться по истории формирования растительного покрова Даурии. Считается, что в плиоцене (9–1 млн. лет назад) по крайней мере в северной части Даурии простиралась широкая полоса неморальных лесов, соединявшая неморальные широколиственные леса Европы и Якутии. Ильмовые харганаты и ильмовые рощи ведут свое происхождение именно от этих третичных лесов, произраставших в гумидную эпоху. Другие остатки этих лесов – черноберезовые рощи на востоке Даурии. По видовому составу они сходны с дальневосточными лесами из дуба монгольского. По всей вероятности, дубовые и ильмовые леса занимали в Даурии в те времена главенствующее положение, но в эпохи последующего похолодания и аридизации "ушли" на восток. В периоды похолоданий, особенно ледниковья, происходило сокращение ареалов широколиственных пород, но полностью они не исчезали, сохраняясь в рефугиумах на южных склонах (Волкова, Белова, 1980). В эпохи похолодания на рубеже межледниковья и ледниковой эпохи и потепления при переходе от оледенения к голоцену наблюдались короткопериодические колебания климата, порождавшие быстрые изменения ландщафтов.
По данным споро-пыльцевых анализов (Евтеева, 1965; Карасев, 2002; Птицын и др., 2010; Решетова и др., 2013; Решетова, 2017), наиболее значимые перестройки состава растительности и структуры ландшафтов в Даурии произошли в позднеледниковье, в раннем и среднем голоцене, когда климат менялся наиболее резко. При этом тундро-степи и лесотундры приледниковых областей сменились степями и лесами. Климат среднего голоцена был более гумидным, а леса гораздо более влажными, чем в настоящее время, и состояли из пихты и ели, что подтверждается наличием их пыльцы в спектрах из отложений многочисленных озер Забайкалья (Решетова, 2017). В период регионального климатического голоценового оптимума вокруг Байкала были распространены елово-кедрово-пихтовые леса, а в северной Даурии шло расширение ареала ельников. Однако уже 5,5–7 тыс. лет назад при нарастании аридности почти все темнохвойные леса трансформировались в светлохвойные – лиственничные и сосновые. Ксеротермический максимум, по оценкам палеографов (Белова 1985), наблюдался в Даурии 5–5,5 тыс. лет назад, когда основным типом растительности здесь стали степи и лесостепи. Именно в этот период сложился особый и специфический комплекс лесостепных растительных сообществ, отличающийся одновременным сосуществованием леса, степи и прерии, между которыми наблюдаются взаимодействие и экотоны разного рода.
Большие дискуссии в свое время вызывал вопрос об устойчивости лесостепи, по которому существуют прямо противоположные мнения. Наиболее предпочтительной выглядит концепция Э.Г. Коломыц (2003), в которой относительно высокоустойчивой признается только северная и центральная подзоны лесостепи, в то время как южная подзона характеризуется крайне низкой устойчивостью и большим динамизмом. Что касается лесостепи Даурии, то растительность ее можно считать крайне неустойчивой, несмотря довольно длительное существование.
Нестабильность растительности Даурии, связанная с постоянными колебаниями климата, порождает неустойчивость видового состава, причем в разных синузиях растительности, по-видимому, скорость изменения этого состава совершенно разная. Так, например, моховая синузия реагирует на колебания климата очень медленно, довольно медленно реагируют лишайники, и очень быстро сосудистые растения. Из последних древесные наиболее медленны, и однолетники наиболее быстры. Можно сказать, колебания в видовом составе травянистых однолетников – это моментальная реакция растительного покрова на годовые климатические флуктуации.
Так как в реальном растительном покрове все синузии контактируют между собой, то при разности скоростей изменения этих синузия возникает некая нестабильность, в какой-то мере аналогичная явлению неустойчивости Кельвина-Гельмгольца в физике. Типичным примером такой физической нестабильности является образование волн на поверхности воды под действием ветра. В растительном покрове под воздействием аридизации или гумидизации тоже образуются своего рода «волны», исходящие из центров климатоформирования региона – центрально-азиатского (пустыни Гоби), восточносибирского (Байкала) и дальневосточного (Тихого океана). Во время "работы" этих волн в растительный покров Даурии поступают порциями виды полупустынь и опустыненных степей, темнохвойной тайги и криофитных степей, дальневосточных широколиственных лесов, манчжурских прерий. В свою очередь, растительность Даурии как пульсирующий центр также "испускает" в растительный покров окружающих ее регионов виды, сформировавшиеся в нестабильных условиях колебания "климатического маятника". В результате взаимодействия этих процессов формируется очень сложная динамическая система растительного покрова данного региона, способная выдерживать любые экстремальные потрясения, предлагаемые ей экосферными оболочками.
В настоящее время одним из "модных" подходов в изучении нестабильности растительности является концепция экотона. Понятие "экотон" претерпело длительную историю, в ходе которой от узкого определения "переходной зоны" трансформировалось в широкую концепцию "экотонных систем" (Якомяги и др., 1988; Соловьева, Розенберг, 2006). В качестве экотона рассматривают переходные, граничные пространства между различными природными системами (экосистемами, ландшафтами), между природными и антропогенными системами, между различными средами и между различными природными зонами (Залетаев, 1997). Появились работы, в которых с точки зрения системного подхода анализируется экотон лесостепь – степь и лесостепь – лес (Малая, 2012; Сизых, 2014). Возникла и терминологическая дискуссия по поводу применения концепции экотонов при рассмотрении разнокачественных явлений – контакта целых зон растительности, контакта природных сред, контакта фитоценозов и т.п. Появилась теория "триггерных" систем как экотонов (Калмыков, 1999; Банникова, 2003), достаточно громоздкая и мало объясняющая суть процессов потери устойчивости экосистем. Пока использование концепции экотонов в геоботанике носит узко-теоретический характер и во многом условно, скорее затрудняя, чем проясняя суть вопроса. Пройдет еще немало лет, пока на этом пути появятся четко разработанные критерии использования этого термина и алгоритм применения теории экотонов к исследованию конкретных пограничных систем. Тем не менее, уже ясно, что анализ границы лес – степь как экотонной парагенетической системы (Мильков, 1966, 1977) имеет свои перспективы. "Парагенетическими" системами называются сопряженные, взаимодействующие друг с другом в пространственном и временном аспекте геосистемы, особенно в зонах контакта. Парагенез можно иными словами назвать инвариантом геосистемы, помогающим делать прогнозные оценки по поводу развития некоего природного явления. Барьерный эффект контактных зон может ослабевать или усиливаться в ходе постоянных изменений среды, что влечет за собой развитие новых ландшафтных процессов. |

Одно из классических экотонных сообществ – светлохвойные (сосновые) саванны, площадь которых тем больше, чем выше амплитуда даурского климатического "маятника". Даурия Селенгинская, Республика Бурятия. Фото с сайта: http://forum.poehali.net/ |
Одно из направлений в изучении лесостепи связано с оценкой баланса влаги в экосистемах. Так, в лесостепи леса приурочены к таким местообитаниям, где частный баланс влаги сдвинут в сторону положительного, остепненные луга – к местообитаниям с нейтральным или слегка положительным частным балансом, луговые степи (даурские прерии) – к местообитаниям с отрицательным частным балансом (Бобровская и др., 2014).
Такая дифференциация экотопов и позволяла многим геоботаникам считать лесостепной ландшафт климатически устойчивым, на котором колебания климата отражаются лишь в количественном соотношении леса и степи, не затрагивая самой сущности лесостепного ландшафта (Мильков, 1951).
В целом следует заметить, что существование системы лесостепи в том виде, в каком мы имеем ее в Даурии, позволяет применять для ее исследования и описания самые разнообразные формально-логические схемы и алгоритмы в зависимости от цели изучения. Разнообразие в этом вопросе приветствуется, если оно базируется не на голых фантазиях, а на неких идеях в понимании истории и развития растительности этой зоны. Несмотря на огромное количество посвященным лесостепям работ, можно сказать, что с динамической точки зрения система лесостепного комплекса Даурии изучена крайне недостаточно. А ведь эта система имеет вполне ясную конструкцию; в ней есть и главные звенья, и узловые моменты, и основные связи.
Позволим себе представить себе эту систему в виде следующей схемы:
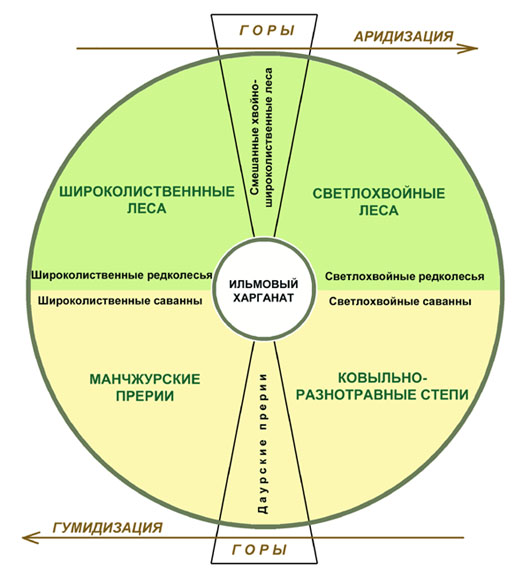
Схема развития лесостепного комплекса растительности Даурии
в системе климатического маятника «гумидизация – аридизация»
|
Здесь Даурия представлена как феномен на границе циклов аридных и гумидных климатических обстановок. В теплые и гумидные эпохи здесь распространены широколиственные леса и прерии манчжурского типа (Васильев, 1944), в прохладные и аридные эпохи – светлохвойная тайга и настоящие степи. Большую часть времени растительность Даурии находится в промежуточном положении, когда присутствуют все элементы, но в разной степени и при разном сдвиге в сторону лесов или безлесья. Так, в современную эпоху граница сдвинута в сторону аридных типов: широколиственные леса представлены лишь фрагментами ильмовых, яблоневых лесов, по горам черноберезовыми рощами; манчжурские прерии уже не наблюдаются и вытеснены на восток за пределы Даурии. Зато в изобилии представлены светлохвойные леса – сосняками и лиственничниками; богато представлены настоящие степи, а на горах – даурские прерии (луговые степи), производные манчжурских высокотравных прерий.
Сочетание широколиственные леса – манчжурские прерии в современную эпоху встречается на Дальнем Востоке России в зоне тихоокеанского муссона. Прерии манчжурского типа занимают Ханкайскую впадину и частично долину Суйфуна (Раздольной) в Приморском крае. Большая часть таких прерий обнаруживается в Китае (Внутренняя Монголия) и частично в Северной Корее.
Типологически пограничными сообществами между степями и лесами в пределах тепло-гумидной климатической обстановки являются ильмовые редколесья и саванны; в пределах прохладно-аридной обстановки – светлохвойные редколесья и саванны. Все четыре экотонных типа хорошо развиты в настоящую эпоху, но в результате элиминации манчжурских прерий, соседствуют либо с даурскими прериями, либо с настоящими степями.
В горных экотопах при переходе одной обстановки в другую формируются два крупных транзитных типа растительности: смешанные (хвойно-широколиственные) леса и даурские прерии (луговые степи). Это типы растительности, несущие в себе черты двух контрастных типов растительности. При этом при аридизации климата на месте светлохвойной тайги образуются в массе остепненные светлохвойные травяные леса, а на месте широколиственных лесов – их фрагменты в степях (ильмовые рощи). В сильно аридные эпохи линия на схеме между лесами и степями сдвигается наверх, в сильно гумидные – опускается вниз, и площади степей и прерий сокращаются.
Горными вариантами остатков широколиственных лесов являются ильмовые харганаты и их производные. На нашей схеме они расположены в центре, так как соединяют в себе одновременно черты лесной и степной формаций. Это предопределяет их большой возраст и относительную устойчивость в постоянно меняющемся ландшафте. |
Благодаря нашей схеме, мы видим в феномене даурской лесостепи сенсорные звенья (Беликович, 2005) – элементы, даже мельчайшее изменения в которых могут индицировать колебания климата и трансформацию всего даурского лесостепного комплекса. Главными сенсорами в этом комплексе являются выделенные нами новые типы растительности – даурские прерии (которые являются скорее лугами с сильным остепнением) и ильмовые харганаты (как отголоски некогда существовавших здесь третичных широколиственных лесов). Оба типа растительности содержат в себе как ксерофиты, так и мезофиты, будучи готовыми приспособиться к любому из состояний климата.
Таким образом, в плане растительности даурская лесостепь функционирует как маятник, раскачивающийся между двумя крайними климатическими обстановками. Этот маятник был не очень заметен в довольно продолжительный период слабых изменений в растительности в середине – конце прошлого века, однако в последнее десятилетие о трансформации даурской растительности стали говорить уже многие ботаники и экологи. Так, в Прибайкальском регионе отмечено потепление и повышение влажности, приводящее к экспансии темнохвойных лесов (Аненхонов,
Кривобоков, 2006; Krivobokov, Anenkhonov, 2012), а в долине Селенги – ксерофитизация степных сообществ (Аненхонов, 2015). Идущие процессы опустынивания стали заметны и для всего Восточноазиатского сектора степей (Гунин, Микляева, 2006).
Что касается центральной и южной части Даурии, то здесь аридизация проявляется наиболее остро. По данным В.А. Обязова (2014), в среднем по территории Забайкалья средняя годовая температура воздуха за период с 1951 по 2010 гг. повысилась на 1,8 °С, а в юго-восточных его районах до 2,7 °С, причем темпы потепления в Даурии значительно (в два с половиной раза) выше, чем в среднем по Северному полушарию. Учтем здесь, что по оценке межправительственной группы экспертов по изменению климата ЮНЕП, тридцатилетний период с 1983 по 2012 гг. был самым теплым в Северном полушарии за последние 1400 лет (!). Рост температуры воздуха приводит к увеличению продолжительности вегетационного периода и смене состава видов в фитоценозах. Наибольшее увеличение вегетационного периода, превысившее 10 дней, произошло в центральных, южных, юго-восточных районах Забайкалья, а также в бассейне р. Селенги. Вероятно, на территории Северной Монголии наблюдается еще большее увеличение этого периода. Неудивительно, что после 2000 г. в Даурии значительно возросло число лесных пожаров, особенно существенно выросла площадь пожаров (чуть ли не в 20 раз, по Обязову, 2010).
В связи с наблюдающимися климатическими перестройками стал широко обсуждаться вопрос о продолжительности и амплитуде ритмов колебания климата, в том числе и Даурии. В Забайкалье выявлены следующие колебания климата: 60-летние, 25–26-летние и 6–7-летние ритмы, имеющие 90%-ную доверительную вероятность (Обязов, 2014). Что касается колебаний в числе выпадаемых осадков, то климатологи затрудняются с выявлением здесь каких-либо трендов, хотя указывают на имеющиеся 30-летние ритмы. В период до 2020-х гг. ожидается повышенный фон атмосферных осадков в середине лета, однако он не будет мешать процессам ксерофитизации растительности. Высохшая почва только смачивается с поверхности идущими дождями, и при росте теплообеспеченности будет продолжаться усиление засушливости. Что же происходит и будет происходить с растительностью Даурии?
Для того, чтобы внятно ответить на этот вопрос, следует обратиться к данным геоботаников прошлого века, исследовавших гидротермический режим в разных зонах даурских и монгольских степей:
Гидротермический режим степей Монголии (по: Береснева, Рачковская, 1978)
Основные типы сообществ |
T1 |
T2 |
Tг |
Период с T>5° |
Осадки, мм |
Луговые степи |
–22,8 |
17,5 |
–1,5 |
135–150 |
250–300 |
Настоящие степи |
–21,8 |
20,0 |
0,2 |
160 |
200–250 |
Сухие степи |
–20,0 |
21,5 |
0,5 |
165 |
150–200 |
Опустыненные степи |
–18,5 |
21,7 |
1,3 |
175 |
120–150 |
Пустынные степи |
–19,0 |
23,0 |
3,4 |
185 |
< 120 |
Примечание: Т1 – среднемноголетняя температура воздуха января; Т2 – то же июля; Тг – то же за год; Период с Т – продолжительность периода (в днях) с температурой воздуха более 5°; Осадки – среднегодовая сумма атмосферных осадков (мм)
Получается, что каждая зона степей отличается от соседней по среднегодовой температуре воздуха всего на один-полтора градуса (а между сухими и настоящими степями всего 0,3 градуса!), а по количеству годовых осадков – на 50 мм. Согласно же приведенным выше оценкам, общее потепление атмосферы в Даурии составило не менее 2,5 градуса, а количество осадков сократилось на 40–100 мм. Это означает, что каждая соседняя зона растительности должна уже была "подвинуть" более северную, если бы не существовало некоей инерции биосферных оболочек. По предположениям специалистов, занимающихся историей растительного покрова, "скорость" изменения растительности при перестройках климата в разных зонах и даже в разных полушариях планеты разная и может составлять от сотни до сотен тысяч лет в зависимости от типа растительности (Хотинский, 1977; Панова, 1982; Красилов, 1997; Изменение климата и ландшафтов ..., 1999; Волкова и др., 2002; Филимонова, 2005; Борисова, 2007; Новенко, 2016; и др.). Растительность отстает от изменений климата на несколько десятков лет, почва – на сотню лет и более.
Несмотря на инерцию растительности относительно климата,
тенденция повышения ксерофитности растительных сообществ стала уже очевидной. Особенно легко она уловима на примере "сенсоров" даурского маятника – даурских прерий и саванн. По данным О.А. Аненхонова (2015), в Селенгинской Даурии наибольшие изменения наблюдаются в составе
настоящих и умеренно влажных луговых степей (прерий). В горах же основным процессом, происходящим при потеплении климата, является сокращение площади криолитозоны и деградация островной мерзлоты. В Хэнтей-Чикойском нагорье за последние 30 лет наблюдаются мощная протайка мерзлоты, полная ликвидация погребенных глетчеров в цирках, ледяных линз в теле морен, резкое потепление воды в озерах и реках верхнего лесного пояса и подгольцового поясов. Южная граница криолитозоны сдвигается на север, идет ее постепенная фрагментация.

Верхняя граница леса особенно чувствительна к климатическим изменениям. Подгольцовые заросли ерника и золотистого рододендрона Rhododendron aureum, с единичными кедрами. Отроги гольца Сохондо, Хэнтей-Чикойское нагорье. Фото: © Татьяна и Игорь Маврины |
Проводимый нами и другими сотрудниками Сохондинского биосферного заповедника долговременный мониторинг растительности на постоянных пробных площадях ((Беликович, 2001, 2005; Беликович, Галанин, 2000, 2002а,б; Беликович и др., 2011а,б; Галанин, Беликович, 1988, 2001, 2002, 2004, 2009а,б; Галанин и др., 2002, 2003, 2007) показал, что в верхнем лесном поясе сокращаются площади моховых и торфяных болот, зеленомошные леса трансформируются в кустарничковые и травяные, сосна вытесняет лиственницу и кедр. В растительном покрове всех типов леса активизируются шиповники, жимолость, сухолюбивые виды осок, но исчезают бореальные кустарнички, сокращается площадь ерников и голубичников. Усиливается контрастность между повышенными дренированными и пониженными заболачивающимися из-за протайки мерзлоты экотопами. Ослабленные засухой и сменами фенологических ритмов, леса становятся легкими жертвами насекомых-вредителей, прежде всего шелкопряда, а исчезновение влажной моховой прослойки и усиление дренированности экотопов приводит к усилению пожароопасности. Возникающие пожары могут приводить к катастрофическим последствиям, так как из верховых становятся низовыми.
Особенно чувствительна к надвигающейся аридизации климата верхняя граница леса в горах (см. фото слева). Заложенные на гольце Сохондо площадки по верхней границе леса показывают, насколько суше стало на нагорных террасах. С большой скоростью идет усыхание кедровостланиковых зарослей и ерников (видимо, из-за снижения снежности и мощности снегового покрова зимой). На некоторых участках наметилась тенденция усыхания и деревьев на верхней границе леса. |
Сами горные дриадовые тундры ксерофитизируются, становятся кобрезиевыми, из их покрова исчезают многие кустарнички. Сокращаются площади осоковых и нивальных моховых тундр. В нижнем лесном поясе идут процессы олуговения и остепнения, языки петрофитных даурских прерий проникают вплоть до подгольцовья. Те же участки горных прерий, что были распространены в нижнем лесном и лесостепном поясе, постепенно видоизменяются в настоящие петрофитные степи. Даурские прерии на надпойменных террасах тоже часто ксерофитизируются и трансформируются в настоящие ковыльно-разнотравные степи. Луга из сырых переходят во влажные или сухие.
Катастрофичность изменений, наблюдаемых нами в южном Забайкалье, весьма впечатляет, но эти выводы контрастируют с осторожными оценками этих процессов со стороны геоботаников, изучавших растительность Западного Забайкалья (Аненхонов, 2015). По всей видимости, на западной границе Даурии климат изменяется медленнее из-за близости мощного фактора – Байкала, мерзлота более инерционна, а мохово-лишайниковая подушка в лесу обладает большими буферными свойствами. Тем не менее, при сохранении тренда потепления климата и здесь можно ожидать начало деградации сообществ таежных мохово-лишайниковых еловых лесов в мерзлотных районах.
Что же будет происходить в зоне степей? Вероятно, ковыльно-разнотравные степи будут вытесняться сухими мелкодерновиннозлаковыми, а последние – "подпираться" опустыненными вариантами степей. Вполне возможны сдвиги между границами растительных зон, которые, к сожалению, трудно фиксировать, так как для этого нет необходимых маркеров и реперов. Но уже сейчас в зоне контакта лесостепей и настоящих степей происходят процессы, которые трудно не заметить: меняется аспект даурских прерий, состав видов и структура ярусов. Требуются специальные мониторинговые работы, чтобы не просто зафиксировать эти изменения, но и понять, в какую сторону "двигается" растительность. Вполне вероятно, что при росте аридности этой субаридной территории может и не произойти упрощение хорологической структуры растительного покрова, как ожидают геоботаники, а наоборот, распространятся мозаичные комплексы, включающие в себя как ксерофильные, так и мезофильные ценозы.
Ясно одно: даже при учете роста осадков вплоть до 2020 г., фоновым процессом станет повышение засушливости, усиляемое ростом частоты лесных и степных пожаров и вспышек насекомых-вредителей леса. Ливневые летние сезоны будут все более напоминать сырые периоды "африканского типа", при которых казалось бы интенсивные дожди и мощные летние паводки приводят только к одному: концентрации жизни в довольно узком временном отрезке и опустыниванию. Думается, что основным сценарием изменения растительности Даурии в ближайщие 50–100 лет станет наступление лесостепей на зону подтаежных лесов, проникновение в зону лесостепей с юга языков настоящих степей, и сдвиг на север границ между ковыльно-разнотравными и сухими мелкодерновиннозлаковыми степями, между сухими и опустыненными степями.
Растительность Даурии на протяжении последних веков и вплоть до настоящего времени – это визитная карточка всех неравновесных растительных систем. На ее примере можно понять законы становления, развития и функционирования экосистем и фитохор, находящихся в зоне контакта контрастных условий и совершенно различных по происхождению и флорогенезу. Динамику состава и структуры растительного покрова Даурии можно объяснить только зная законы и амплитуду климатического "маятника", раскачивающегося над территорией юга Восточной Сибири и Центральной Азии. Понимая импульсы условий среды, связанные с климатическими и антропогенными изменениями, и время этой нестабильности, можно прогнозировать векторы во флористическом и хорологическом разнообразии растительности, а уже исходя из этого, оптимизировать природопользование в этом уникальном регионе. Именно поэтому исследования фенологических ритмов и мониторинг экосистем для выявления трендов климата, растительности и в целом природной обстановки становятся первоочередной задачей всех научно-исследовательских институтов и природоохранных организаций региона, особенно биосферных заповедников.
