Вы находитесь: НАША БОТАНИЧКА — Растительность — РАСТИТЕЛЬНЫЙ ПОКРОВ СЕВЕРО-ВОСТОКА РОССИИ
РАСТИТЕЛЬНЫЙ ПОКРОВ СЕВЕРО-ВОСТОКА РОССИИ
А.В. Галанин, А.В. Беликович © 1996
Динамика растительности (хронологический аспект растительного покрова)
Растительный покров – равно его флора и растительность – на
любом участке земной поверхности не остается неизменным. Наряду с длительными
изменениями, которые совершаются в геологических масштабах времени, мы часто
сталкиваемся с так называемыми растительными сукцессиями – изменениями, которые
происходят в экологическом масштабе времени (десятки сотни лет). Как известно,
факторами динамики флоры и растительности любого ландшафта могут быть
экзогенные изменения рельефа, изменения климата, литогенной основы, атмосферы,
тектоники литосферных плит, морские трансгрессии и регрессии, надземные и
подземные оледенения, деятельность вулканов, процессы видообразования, пожары,
наводнения и антропогенные факторы. В нашем регионе все эти факторы активно
влияли и влияют на растительный покров. Одни из них вызывают исторические смены
в геологическом времени, другие обусловливают изменения в экологическом времени – сукцессии.
Под воздействием внешних факторов устойчивый растительный
покров нарушается, локальные экосистемы выводятся из состояния
биогеохимического равновесия, в них образуются свободные экологические ниши,
поселяются новые виды и жизненные формы, нарушается равновесие в конкурентных и
аллелопатических отношениях между растениями – и все это, в конечном счете, реализуется
в виде заметных и весьма резких изменений ассоциаций, синузий и фитоценозов.
Если затем действие фактора приостанавливается (экологический режим
стабилизируется), растительный покров через серию смен возвращается в близкое к
исходному состоянию.
Основные типы сукцессий
1. Сукцессии в долинах рек. Легче всего пронаблюдать динамику
растительности в долинах рек Пойменные сукцессии представлены здесь
многочисленными сообществами, изменяющимися в пространстве по мере удаления от
русла реки, и могут рассматриваться как временные ряды растительных сообществ.
Благодаря большой скорости аллювиальных процессов, сравнимой с человеческой
жизнью, эти сукцессии легко улавливаются и хорошо изучены геоботаниками и
лесоведами. Это естественные смены сообществ по ходу увеличения возраста вида-доминанта
и по мере ослабления аллювиального процесса.
По всем крупным рекам региона выделяется 3–4 уровня поймы.
Поймой считаются все земли, заливаемые паводковыми водами хотя бы раз в
несколько лет и испытывающие в связи с этим влияние потока вещества и энергии,
несущегося рекой. Это влияние может сказываться в отепляющем действии реки
часто под руслом находятся таликовые зоны, распространяющиеся и на пойму, либо
в наносе большого количества гальки, песка, иловых частиц, плавника. Пойменные
сообщества служат как бы преградой на пути смыва вещества рекой и работают
против потока, усиливая аккумуляцию. Так, все растущие здесь растения обладают
способностью в большей или меньшей степени задерживать влекомые мутными водами
мусор и опавшие листья. Таковы ива, тополь, но более всего эта способность
развита у чозении, которая первая поселяется на промытых галечниках и по мере
роста начинает быстро формировать "щетку" из торчащих боковых
прутьев. Эта своеобразная "юбка" задерживает несущийся материал,
накапливает мелкозем, образуя первичный почвенный слой, способствующий
поселению здесь других растений. Чозения по праву считается королевой поймы
наших северных рек она формирует ее и дает начало пойменной сукцессии.
По мере отступания русла и с появлением мерзлоты чозения
выпадает из сообщества, сменяясь лиственницей. На галечниках Арктики подобную
лиственнице роль выполняет ива аляскинская. Река может иметь несколько
надпойменных террас, однако растительность на них изменяется гораздо медленнее,
чем в низкой пойме, и генезис этих смен уже не связан с рекой.
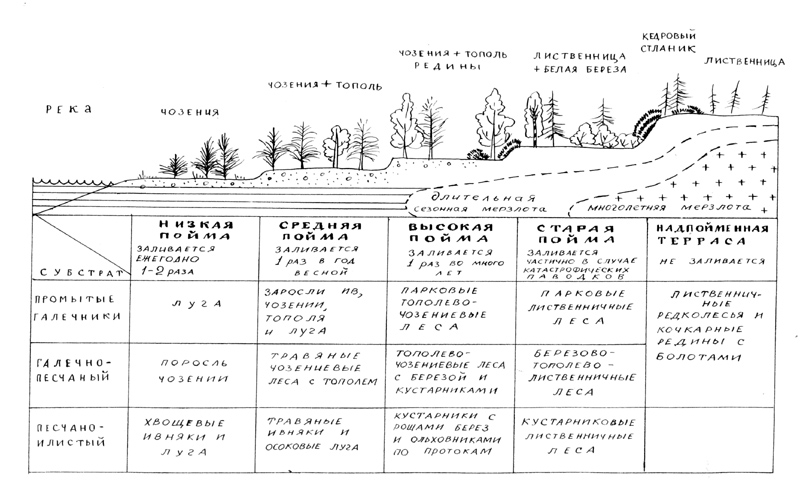
Рис. 11.
Схема типичного распределения пойменной растительности по уровням поймы
|
Типичная
пойменная сукцессия показана на рисунке слева (рис. 11). Подобное распределение
сообществ характерно для крупных лесных рек региона – Омолона, обоих Анюев и их
основных притоков, верхнего течения р. Анадырь. На небольших речках сукцессия
может иметь менее развитый характер, однако последовательность смен остается
той же.
Сукцессия может
отсутствовать или может иметь некоторые искажения на наледных участках долин,
где растительность приспосабливается к условиям периодического подтопления и жесткого
оледенения. Чаще всего наледь как бы выравнивает пойму, делая дно долины
плоским, и разница между уровнями поймы исчезает. Все дно долины в этом случае
занимает один комплекс растительности – низких, как бы
"подстриженных" морозом ивнячков, голубичников и кочкарных лугов, и
лишь вдоль многочисленных русел реки в зоне наледеобразования наблюдаются фрагменты
растительности низкой поймы. Кроме того, из-за неустойчивости русла, которое в
наледной долине свободно "гуляет" по долине, наблюдается множество
параллельных проток, растительные сообщества здесь несут на себе печать
временности. Древесные растения не успевают закрепиться по берегам проток, а луга
не развиваются из-за периодического заливания, старые же протоки нередко
заболачиваются. |
На участках мощных наледей вся долина реки может быть занята
почти голыми галечниками с куртинами злаков и разнотравья. На крупных наледях
запасы льда бывают столь велики, что не успевают растаивать в течение лета. В
Магаданской области самые мощные наледи (до 4–6 м толщиной и до 10–15 км длиной)
развиты на р. Анмагында (приток р. Детрин), Лабазная (приток Омолон), Верхний
Орах (приток р. Кегали). На Чукотке такие наледи вполне обычны. Окраины
гигантских речных наледей в лесной зоне вне Арктики часто являются
местообитанием для многих арктических видов.
2. Зарастания отвалов зодотодо6ычи. Золотодобывающая
промышленность вызывает в нашем регионе наиболее крупномасштабные нарушения
растительного покрова. Всего, по данным статистики, в регионе нарушено свыше
135 тыс. га земель, около 30 тыс. га - на Чукотке. Это в основном долины рек и
ручьев, до золотодобычи занятые наиболее продуктивными в ландшафтах пойменными
лесами, кустарниками и болотами.
Масштаб нарушений и изменение энергетики техногенного
ландшафта (а геофизические исследования показали значительное возрастание
тепловых ресурсов нарушенных земель по сравнению с естественными) подтолкнули
северян к использованию этих земель в сельскохозяйственных целях. В 1970–1980-е
годы делались успешные попытки создать на распланированных отвалах
искусственные луга, а также поля кормовых и овощных культур. К сожалению, из-за
глубокого кризиса эти работы сейчас приостановлены в связи с их экономической
нерентабельностью. В итоге на сегодняшний день лишь ничтожная часть земель
(менее 0,01%) в регионе рекультивирована. С 1992 г. в связи с общим упадком в
золотой промышленности все работы по рекультивации фактически прекращены.
Открытая разработка россыпных месторождений золота ведется
двумя способами – карьерным и дражным. При обоих способах разработки весь
растительный покров сдирается, и вся толща непродуктивных по золоту пород
снимается (так называемая "вскрыша торфов") и складируется в
специальных вскрышных отвалах. Выемка пород производится бульдозерами,
экскаваторами, самосвалами, перед этим породы оттаивают различными способами.
Вскрышные отвалы имеют естественный состав грунтов смесь торфа, мелкозема,
щебня, валунов, почвы, и поэтому наиболее благоприятны для зарастания
растениями.
После удаления так называемых "торфов" образуется
голый полигон продуктивных по золоту пород (так называемые "пески").
При мелкокарьерном способе разработки они снимаются бульдозерами и обогащаются
на промывочных приборах (промприборах). В процессе обогащения эти породы
разделяются на крупнофракционные (щебень, галька, гравий, валуны), образующие
"галечные" отвалы, и мелкофракционные (песок, дресва, глины), которые
сливаются в виде пульпы в гидравлические, так называемые "эфельные"
отвалы. Кроме эфелей, отходы обогащения поступают в хвостохранилища и илоотстойники.
При дражном способе разработки выемка "песков" и
их обогащение происходят на драге, которая медленно движется по оттаянным
породам и выбрасывает через кормовые колоды эфельный материал, поверх которого
выкладывает хорошо промытые крупнофракционные "галечные" отвалы.
Между дражными отвалами и бортами котлована остаются заполненные загрязненной
водой пазухи. В результате формируется новый техногенный ландшафт, состоящий из
отвалов и пазух. Растительность, появляющаяся на этих землях первой, зависит от
способа формирования отвалов.
Дражные галечные отвалы зарастают медленнее всех остальных
отвалов, и зарастание идет вилами галечных кос и пойменных наносов – злаками
(Elymus sibiricus, Trisetum molle и др.), звездчаткой Фишера (Stellaria
fischeriana), астрагалами Astragalus schelichovii, Astragalus alpinus и т.д.
Здесь хорошо возобновляются чозения, тополь и все виды пойменных ив: Salix roridа,
Salix udensis, Salix schwerinii. Обычны виды, предпочитающие хорошо прогреваемые
галечники скерда малая (Crepis nаnа), прострел даурский (Pulsatilla davurica), горошек
Vicia multicaulis, остролодочник Oxytropis vasskovskii и др. Сукцессия здесь
направлена на восстановление тополево-чозениевых лесов. По мере удаления от
русла реки с течением времени может формироваться долинный лиственничный лес,
постепенно заболачивающийся. Вероятно, через 400–500 лет комплекс заболоченных
кустарников и лиственничного редколесья, характерный для широких долин Bepxнeколымского
округа, может восстановиться полностью.
Bскpышныe отвалы – автотранспортные, а также скреперные,
бульдозерные, гидравлические (шахтные смывы, хвостохранилища и илоотстойники) –
зарастают, как и разведходы и вырубки, в основном сорными, заносными видами с
широкой экологической амплитудой – такими, как дескурения, сурепка, вейник
Лангсдорфа, иван-чай узколистный, жерушники (Rorippa barbareifolia, Rorippa
palustris), желтушник Еrysimum cheiranthoides, донтостемон Dontostemon ресtinatus,
скерда кровельная (Crepis tectorum), мелколепестник Erigeron politus, пукчинелла
Puccinellia hauptiana. В большинстве случаев сукцессия здесь направлена на
восстановление лиственничных лесов. Естественной растительностью быстрее всего
зарастают отвалы вскрыши, так как они часто содержат в себе покоящиеся семена,
и их состав наиболее благоприятен для формирования первичной почвы.
Довольно своеобразным местообитанием для вновь поселяющихся
растений являются высыхающие илоотстойники с суглинистым материалом. Здесь
поселяются иван-чай узколистный, лютик ядовитый (Ranunculus sceleratus), крестовник
арктический (Senecio congestus), лисохвост Alopecurus aequalis, бекмания Весkmanniа
sizygachne, хвощ луговой (Equisetum pratense), лужница водная (Limosellа aquatica),
нардосмия холодная (Nardosmia frigida). В дальнейшем дно илоотстойника либо
заболачивается, либо высыхает, и его суглинистая поверхность растрескивается.
|
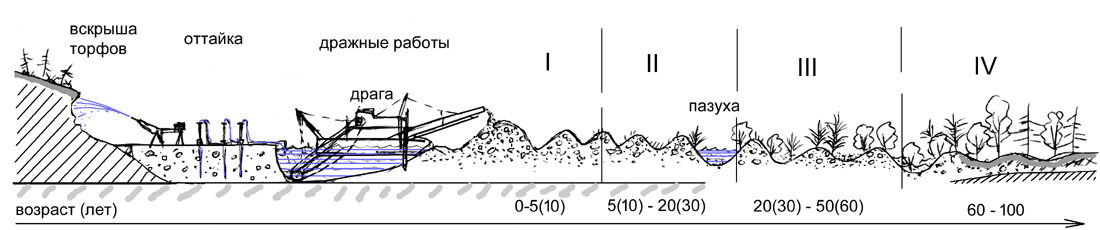
Типичный ход процесса зарастания на дражных отвалах показан
на рис. 12.
Рис. 12. Сукцессии растительности при добыче россыпного золота на землях, нарушенных золотодобычей в долине реки (на дражных отвалах). Вверху: последовательность операций при добыче россыпного золота и сукцессия растительности в пойме. Внизу: сукцессия растительности на надпойменных террасах. Этапы (фазы) сукцессии:
I – отсутствие растительности;
II – пионерная стадия (редкотравные луга, возобновление чозении, открытые группировки)
III – ивняки с чозенией
IV – ивово-лиственничный лес
V – лиственничный лес
VI – заболачивающийся лиственничник |
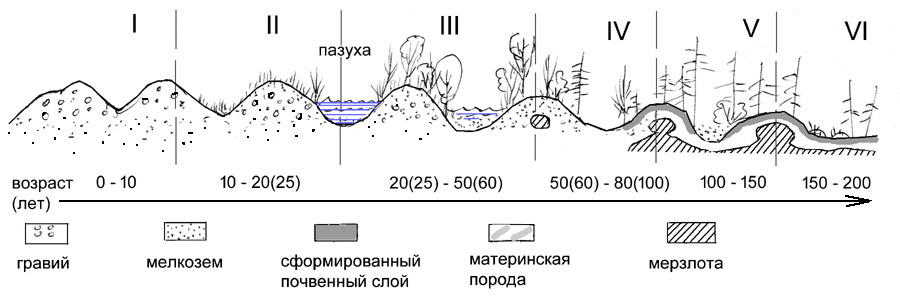 |
Нулевая фаза характеризуется отсутствием растений. Она длится 2–5
лет после образования отвала, пока идет миграция диазачатков (семян, спор) с
соседних уже заросших участков либо с участков, где был сохранен естественный
растительный покров. Надо сказать, что количество выпадающих на поверхность
отвала диазачатков (интенсивность семенного дождя) – очень важный фактор
скорости зарастания и видового состава пионерной растительности. Первая фаза
сукцессии – заселение видами с легкопереносимыми семенами. Часть видов не
выносит условий жизни на обнаженном промытом грунте и вскоре гибнет. Остаются
виды, обладающие высокой активностью захвата территории (многие сорные), либо
виды местной флоры, приспособленные расти на галечниках. Постепенно происходит
обогащение пионерного сообщества новыми видами и жизненными формами –
появляются кустарники, деревья.
В настоящее время этот процесс достиг II–III стадий лишь на
участках, начатых отрабатываться в 1930–1940-е годы. При наиболее оптимальных
эдафических условиях ивняки на отвалах могут появиться уже через 15–20 лет
после нарушения, а через 30-40 лет такие сообщества уже неплохо ценотически
сформированы и можно говорить об их устойчивом длительном существовании, пока
развивается синузия лиственницы. Под пологом лиственницы начинают формироваться
синузии зеленых мхов, гипоарктических кустарничков и кедрового стланика,
характерные для коренной климаксовой растительности региона.
Таким образом, в целом характер и скорость зарастания
нарушенных земель зависит от наличия близких источников семенного материала
оставленных в пойме "островов" с кустарниками и чозениевыми лесами,
не пройденных драгой; в долинах ручьев от соседства склоновых лесов. Если же
при разработке долины ручья на бортах долины лес был сожжен или вырублен,
сукцессия на отвалах может замедлиться на 40–60 лет. Поэтому для ускорения
процессов самозарастания при дражных работах рекомендуется оставлять небольшие
участки (10–50 кв. м) с нетронутой растительностью, которые могут
разрабатываться позже при повторном прохождении.
К этим наиболее дешевым способам ускорения самозарастания
отвалов можно еще добавить гидропосев семенами трав, кустарников и деревьев с
дорог и с воздуха Одним из наиболее сложных объектов рекультивации являются
дражные отвалы, так как технология добычи золота посредством драги предусматривает
насыпку гравийно-галечного материала поверх мелкофракционного эфельного, что
абсолютно неэкологично и неэкономично, если затраты на рекультивацию включать в
стоимость золота.
Существующие в мировом опыте методы предусматривают снятие
почвы перед отработкой долины и ее складирование за пределами зоны разработок,
а затем ее повторное использование или же нанесение поверх дражных отвалов
вскрышных пород. Однако в нашем регионе все эти мероприятия не выполняются с целью
снижения себестоимости добычи золота. Только плата за землю и биологические
ресурсы при всех видах землепользования может стимулировать горняков проводить
рекультивационные работы.
3. Послепожарные сукцессии. Пожары в лесных экосистемах
являются естественным фактором, вызывающим периодический быстрый перевод
накопленной живой и мертвой органики в минеральные вещества. Сгораемая органика
представляет собой древесину, мхи и лишайники, подстилку, отчасти гумусовый
слой, разлагающиеся в норме очень медленно – за 400–500 лет. Всю эту органику
можно рассматривать как резерв питательных веществ для растений, резко
высвобождающийся только после пожара. В течение нескольких лет гарь
представляет собой место с повышенной трофностью, своего рода биогеохимическую
аномалию. Высвобожденные минеральные вещества довольно долго сохраняются на
равнинных, долинных участках, где смыв идет медленно. В то же время на крутых
склонах гор пожар вызывает мощную потерю экосистемой общего запаса азота,
переводя его в катастрофически быстро выносимую минерализованную форму.
Обогащение почвы азотом вызывает в первые после пожара годы
быстрый рост травянистых растений, причем первыми и наиболее массовыми почти
всегда бывает иван-чай узколистный. Затем появляются вейники – Саlamagrоstis
lapponica, Calamagrostis langsdorffii, другие злаки и разнотравье. Пышная
травянистая растительность, питающаяся легко усвояемыми минеральными
веществами, с течением времени (10–12 лет) начинает замещаться различными
кустарниками, мхами, лишайниками, что замедляет процессы вымывания химических
элементов как с поверхности, так и из почвенного профиля.
В целом можно сказать, что пожар резко интенсифицирует
круговорот вещества в экосистеме, ускоряет обращение атомов в петле жизни и тем
самым играет положительную биогеохимическую роль в ландшафтах. Однако в горных
районах на крутых склонах смыв вещества после пожара происходит настолько
интенсивно, что ни о каком сколь-либо быстром восстановлении растительности, а
тем более почвы говорить не приходится. Пожары в горах вызывают мощную эрозию,
таяние мерзлоты и понижение верхней границы многолетнемерзлого слоя сукцессия
на некрутых склонах идет в 2–3 раза медленнее, чем на равнинных участках, а на крутых
склонах она затягивается на многие столетия.
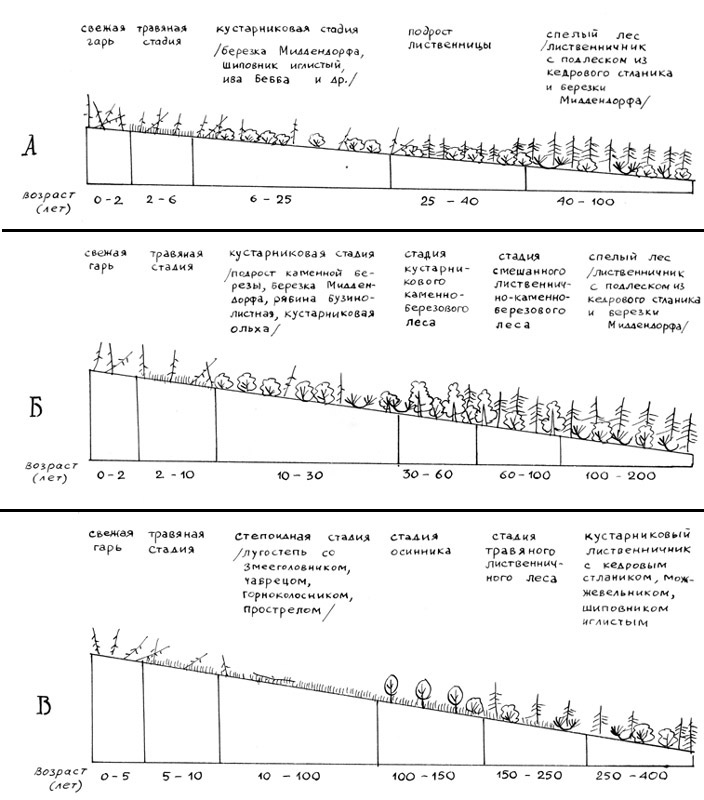
|
Схемы разных вариантов послепожарных сукцессий приведены на
рис. 13.
Рис. 13. Разные варианты послепожарных сукцессий лиственничных лесов Северо-Востока России: А – на склонах и шлейфах склонов (сукцессия по кустарниковоберезовкому типу); Б – на склонах приюжных экспозиций в прибрежных районах Охотского моря (сукцессия по каменноберезовому тьипу); В – на южных склонах в резко континентальных районах (сукцессия по степоидно-осиновому типу). Возраст стадий дан ориентировочно.
Так или иначе, но во всех вариантах послепожарной сукцесии первая
стадия – это мощное зарастание травами. Затем зарастание может пойти по
березовому или осиновому типу, то есть при восстановлении сгоревших лиственнични
ков экосистема обязательно проходит через стадию мелколиственных лесов или
кустарников.
На Охотском побережье широко распространена послепожарная
сукцессия с промежуточной стадией в виде каменноберезового леса. По Колыме
вместо каменной березы чаще можно встретить березу плосколистную. Как в
Приохотье, так и на Колыме сукцессия нередко идет через стадию кустарниковых
зарослей из дальневосточной березки и березки Миддендорфа. В местообитаниях с
повышенной трофностью вместо зарослей березки на этой стадии может развиваться
синузия кустарниковой ольхи.
Гораздо реже и на олиготрофных участках можно встретить
сукцессию, в которой после сгоревшего лиственничника формируются заросли
кедрового стланика – это очень медленная сукцессия, в которой восстановление
лиственницы в ближайшие 4–5 десятилетий весьма сомнительно. Обычно по этому
типу сукцессия идет на крутых склонах в многоснежных районах, часто приморских.
В резко континентальных районах на крутых южных склонах гор как стадия после
пожара часто появляются рощи осины В бассейне р. Челомджа осиновые леса как
послепожарная стадия играют одну из ведущих ролей в ландшафте, замещаясь со
временем на лиственнично-березовые и лиственничные. В бассейне р. Колыма на
южных крутых склонах гор перед стадией осинника в сукцессии могут идти
остепненные ксероморфные луга, при этом они могут оказаться самой
продолжительной стадией в сукцессии – до 100–150 лет и более. |
Иногда гари успешно заселяются лиственницей с самых первых
стадий уже на 3–5-й год после пожара. В этом случае всходы лиственницы
появляются уже в течение травяной стадии, переходя в подрост раньше, чем
произойдет поселение кустарниковых березок или кедрового стланика. Особенно
быстро и почти сплошь заселяется лиственницей на второй стадии зарастания гарь,
возникшая под ее семенной год. Здесь интенсивный семенной дождь оказывается
мощным фактором сукцессии.
В тундровых районах послепожарные сукцессии проходят совсем
по-другому, чем в тайге, и вызывают несколько другие эффекты. В связи с отсутствием
сколь-либо существенного количества быстровоспламеняющегося вещества в тундре
пожары часто носят поверхностный характер. После тундрового пожара в кочкарных
тундрах в центре кочек сохраняются живые растения, почки возобновления, узлы
кущения. Поскольку в кочкарной тундре преобладают растения с вегетативным
размножением, то самовосстановление нарушенных участков происходит сравнительно
быстро – за 10–12 лет. Этому способствуют условия, складывающиеся после пожара –
возрастает глубина протаивания почв, что вызывает их обводнение; повышается
аэрированность почв и содержание в них минеральных веществ. В целом, конечно,
сукцессия в тундре идет в определенной зависимости от характера исходных
сообществ, от типа местообитания. В кустарничковых тундрах пионерными
растениями являются зеленые мхи, вейники, осоки. Пирогенная тундра быстро
увеличивает свою продуктивность за счет сильного разрастания пушицы влагалищной
из сохранивmихся узлов кущения, арктополевицы, вейников. При этом на первой
стадии выпадают почти все виды кустарничков – они обычно полностью сгорают. На
второй стадии вейники и другие злаки постепенно снижают свое обилие, исчезают,
но восстанавливается березка тощая, а за ней и другие кустарнички. Через 8–20 лет
сообщество может приобрести близкий к исходному облик.
Медленнее и неравномернее идет сукцессия в осоково-пушицевых
тундрах с полигонально-жильныии льдами, где после пожара образуются
термокарстовые просадки, озерки, на их склончиках развиваются овраги,
усиливается плоскостной смыв. По данным Б.А. Павлова, на третий год после пожара в таких
сообществах появляется маршанция многообразная, практически отсутствующая в
естественных тундрах этого типа. Она вытесняется мхами из рода Polytrichum
только на 7-й год. В целом характер восстановления идет тем же путем, что и в
мохово-кустарничковых тундрах через вейниковую стадию – однако нарушенный
фитоценоз здесь полностью не восстанавливается, а приобретает новые черты.
Особенно опасны пожары в сухих лишайниковых тундрах на склонах и нагорных
террасах, где лишайниковый покров не восстанавливается полностью в течение
нескольких столетий. В сильно засушливые годы пожары могут охватить и торфяные
тундровые болота, при этом при полном выгорании торфяника исходная экосистема
может никогда не восстановиться, а превратиться в другой тип тундр. Так,
известны случаи, когда в результате развития процессов пучения и заболачивания
на месте сгоревших бугорковатых кустарничковвх и кустарниковых тундр образовывались
полигональные осоково-пушицевые тундры. Вообще сроки и типы пирогенных смен в
растительности в тундровой зоне изучены очень слабо. В кедровостланиковых тундрах
Пенжинско-Среднеанадырского округа послепожарные сообщества можно часто узнать
по доминированию ивы сухолюбивой (Salix bebbiana) и разрастанию арктополевицы
(Arctagrostis latifolia).
4. Сукцессия на термокарстовых озерах. В тундровой зоне
нашего региона можно часто встретить осушенные термокарстовые озера, прорванные
в реку, море или соседнее озеро, располагающееся на более низком уровне. На дне
такого осушенного озера на I стадии сукцессии формируются арктофиловые луга (Arctophila
fulva, Senecio congestus, Caltha arctica, Dupontia psylosantha и др.) высокой продуктивности,
так как верхние горизонты озерных илов достаточно плодородны. Через 2–3 года
эти сообщества сменяются злаковыми и злаково-разнотравными лугами (вейники Calamagrostis holmii, Calamagrostis neglecta, Сalamagrostis purpurea; лисохвосты Alopecurus aequalis, Аlopecurus glaucus; мятлики Роа alpigena, Рoa alpina и
др.). В этой короткой стадии (1–2 года) днища озер являются кормовыми угодьями
для многих птиц и зверей. Затем на поверхности начинается формирование клиньев
полигонально-жильных льдов, поверхность становится бугристой, появляются
арктополевица, гипоарктические кустарнички, зеленые мхи, осоки (Саrех lugens),
пушицы (Eriophorum vaginаtum, Eriophorum polystachyon). Луга отундровевают. В
завершение сукцессии здесь формируется бугорковатая кустарничково-осоковая
тундра, затем развивается полигонально-валиковый микрорельеф, котловина заболачивается
и образуется типичная полигональная осоково-пушицевая тундра. Мерзлотные
условия полностью восстанавливаются довольно быстро – меньше, чем за 10–15 лет.
Подобное же "олуговение" можно наблюдать в тундре
всюду, где идет разрушение почвенно-растительного покрова – при пожарах,
строительстве, транспортных нарушениях и т.п., вызывающих глубокое протаивание
сильнольдистой мерзлоты. Проезд летом любого гусеничного транспорта (вездеходов,
тракторов) вызывает уплотнение и разрыв дернины, что, в свою очередь, ведет к
развитию процессов термокарста и заболачиванию Многократное использование одной
колеи машинами становится невозможным, и трасса обычно расширяется на десятки,
а то и сотни метров. Сукцессия, возникающая после прекращения воздействия
транспорта на тундру, длится от 5 до 40 лет в зависимости от степени нарушения,
характера залегания многолетней мерзлоты и ее льдистости, в зависимости от типа
исходных растительных сообществ. В первые годы появляются вейники Лангсдорфа и
лапландский, мятлики, нардосмия холодная, через 4–5 лет интенсивно включаются
кустарнички (брусника, шикша, березка тощая, ивки) и зеленые мхи.
Схожие эффекты олуговения вызывают зоогенные нарушения – например,
сусликовины, песцовые норы и порои, которые обычно легко индицируются в
ландшафте пышной растительностью. Здесь можно встретить лютик родственный
(Ranunculus affinis), живокость Шамиссо (Delphinium chamissonis), незабудку Мyosotis asiatica, аконит анадырский (Aconitum productum), копеечник
копеечниковидный (Нedysarum hedysaroides), астрагал зонтичный (Astragalus
umbellatus), иван-чай узколистный, арктополевицу, вейники и др., на Восточной
Чукотке в составе таких луговин обязательно встречается горечавочка близкая
(Gentianella propinqua).
Существование в
естественных тундрах лугов и луговой растительности неоднократно привлекало
внимание специалистов сельского хозяйства. Однако кратковременность стадий
олуговения не позволяет широко использовать этот тип сообществ. В 1969 г. появилась
остроумная разработка С.В. Томирдиаро по искусственному управлению
озерно-термокарстовым процессом для получения стабильных лугов. Для этого он
рекомендовал периодически затапливать котловины ранее спущенных озер на 3–4
года для частичного восстановления подозерного талика, и чередовать периоды
затопления и эксплуатации под луга. Однако на практике эта разработка стала
внедряться половинчато в виде одноразового спуска озер, что привело к ликвидации
ряда локальных озерных экосистем, и в конечном счете – к дискредитации идеи,
так как в связи с быстрым заболачиванием вновь возникших лугов затраты на их
создание себя не окупили.
5. Сукцессия сбитых оленьих пастбищ. Общая площадь оленьих
пастбищ в регионе достигает 1,2 млн. кв. км, однако с каждым годом площадь их
снижается из-за промышленного освоения, пожаров и повреждения транспортом за
прошедшие 30 лет территория кормовых угодий сократилась на 67 тыс. кв. км, то есть на 11 % изначальной площади, а оленеемкость при этом уменьшилась на 88 тыс.
голов, или на 11% поголовья.
Из-за завышения плановых, показателей по расчетной
оленеемкости пастбищ и величины оленьего стада после 1969 г. пастбища стали
быстро деградировать, особенно зимние (ягельники), наиболее уязвимые при
перевыпасе. В связи с неупорядоченным выпасом оленей и пере грузкой пастбищ
сокращение запасов ягеля происходит со скоростью 2,5–3,0% от его общего
количества в год. Восстановление же потравленных пастбищ происходит в течение
10–15 лет, что связано с крайне медленным отрастанием кустистых лишайников (по
1,7–6,0 мм в год). Сукцессия здесь идет аналогично смене растительных сообществ
при транспортных нарушениях – через стадию злаков, затем активизируются зеленые
мхи и осоки. Кустарнички, и особенно лишайники, восстанавливаются в последнюю
очередь.
Развитие растительного покрова в плиоцен-плейстоценовое время
и в голоцене
В конце плиоцена
(примерно 2 млн. лет назад) на всей Земле произошло резкое похолодание,
результатом которого стала смена флористического комплекса
тургайских листопадных широколиственных лесов сначала комплексом темнохвойной,
а затем и светлохвойной тайги. По-видимому, в это же время на самом севере
Берингии (мосте суши, соединяющем Восточную Азию и северную Америку)
происходило формирование и безлесных тундровых ландшафтов отчасти с помощью
видов-выходцeв темнохвойной и светлохвойной тайги, а отчасти видов, спускавшихся
на равнины осушенного шельфа арктических морей из альпийского пояса гор Аляски
и Чукотки.
В эпохи морских
регрессий (понижение уровня моря) Ледовитый океан в течение четвертичного
периода неоднократно оказывался замкнутым в окрестностях Северного полюса единой
циркумполярной сушей. Поэтому периоды существования Берингии как обширного
сухопутного моста это всегда периоды похолодания в ее северной и центральной
частях и потепления в южной пацифической части. Климат в обширной осушенной
Берингии был весьма континентальный и холодным, только на юге в эти эпохи выпадало
много осадков и развивались горные и горно-долинные ледники. По всей
вероятности, северная часть Берингийского моста была тундровой, средняя тундрово-степной
и лесостепной, а по южной кромке (во всяком случае, в конце плиоцена) росли
темнохвойные еловые леса, подобные тем, что сегодня растут на Аляске. Охотская
и Камчатская темнохвойная тайга через массивы еловых лесов вдоль Пенжины,
Анадыря и Корякии и южную часть Берингийского моста в эпохи морских регрессий в
плиоцене еще сообщалась с темнохвойной тайгой Аляски. Однако уже в начале
плейстоцена (примерно 1 млн. лет назад) этот флористический канал миграции видов сильно ослаб и в конце
концов оборвался. В это время и в южной части периодически появлявшегося в иллинойсе
Берингийского моста суши темнохвойные леса не восстанавливались на всем
протяжении. Вероятно, здесь в ту эпоху преобладали безлесные
лесо-лугово-6олотные ландшафты с доминированием березы, сосны и лиственницы.
В историческом плане остается неясным, почему столь обычный
в северо-Восточной Азии вид океанического склада, как кедровый стланик,
отсутствует в Америке. По крайней мере, этот факт говорит о том, что условия
Берингийского моста в любой его части в плейстоцене были неблагоприятны для
расселения этого вида. Таковыми могли быть либо суровость и континентальность
климата, либо жесткая конкуренция со стороны ели и пихты. При морских
регрессиях по мере образования моста суши Берингия заселялась уже сложившимися
комплексами и синузиями как со стороны Аляски, так и со стороны Азии. Где-то в
средней части моста эти комплексы встречались и взаимопроникали друг в друга,
одновременно конкурируя, так как друг для друга они являлись ценотическими
барьерами. Похоже, что в моменты таких контактов азиатские ценотические системы
оказывались более сложившимися, чем аляскинские, и сильнее препятствовали
проникновению видов из Америки в Азию. В среднем плейстоцене мощная морская трансгрессия,
при которой Камчатка с Корякией превратились в остров, была континентальных
районах [17].
и его гуиидиэацией в ранее Темнохвойные еловые, сопряжена с
потеплением климата елово-березовые, лиственнично-еловые леса и заросли
кедрового стланика в это время, вероятно, отступали вслед за морской
трансгрессией и образовывали полосу вдоль Охотского моря вплоть до основания
Чукотского полуострова, а кедровый стланик с кустарниковой ольхой, по-видимому,
заходили и на сам Чукотский полуостров.
В конце плейстоцена произошла новая крупная регрессия и висконсинское
оледенение альпийского типа в южной и центральной частях Берингийской суши. От
темнохвойной тайги остались отдельные реликтовые массивы в долине р. Камчатка,
в среднем течении р. Анадырь, вероятно, по р. Майн и вдоль Охотского
побережья. В эту эпоху начали формироваться редкостойные лиственничники современного типа с подлеском из
кедрового стланика и березки Миддендорфа. Ландшафты Берингии в то время в
северной и средней частях были тундровыми и тундростепными, а в южной части шла
полоса кустарников (в основном ивняков из ивы шерстистой, ивы аляскинской) и
лугов, перемежавшихся с участками безлесных болот. Даже в долинах рек Берингии
того времени лесная растительность развивалась плохо, о чем свидетельствует
отсутствие на Аляске столь характерного для Северо-Востока России вида, как
чозения. Южные склоны, дренированные террасы и водоразделы в то время были
заняты криоксерофитными сообществами с доминированием осок и кобрезий (Саrех
obtusata, Саrех rupestris, Саrех supina subsp. spaniocarpa, Саrех duriuscula, Кobresia
bellardii). Здесь весьма обильными были Artemisia frigida, Selaginella selaginoides,
Helictotrichon krylovii, Pulsatilla multifida, Роа glauca, Festuca lenensis, Festuca
anriculata, Festuca altaica [2]. Мы не имеем права называть эти сообщества степями в
принятон смысле этого термина. В современном растительном покрове эти сообщества
не сохранились, хотя отдельные их компоненты – виды и ценоэлементы – мы можем
наблюдать и на Колыме, и на Чукотке, и даже на Охотском побережье.
Последняя фаза
существования Берингийского моста суши закончилась в самом конце плейстоцена
примерно 12–15 тыс. лет назад. Новая трансгрессия, конечно, началась
значительно раньше и постепенно отвоевывала у Берингии все новые и новые пространства
как со стороны Пацифики, так и со стороны Арктического бассейна. Мост суши
истончался, климат на нем становился все более гумидным – более теплым зимой и
более холодным летом. Лесная и лесотундровая растительность южной Берингии в
эпоху узкого моста деградировала, частично отступая на Колыму и вглубь Аляски.
По мере
трансгрессии быстро деградировали и ксерофитные сообщества центральных районов
Берингийского моста, уступив место на Чукотке тундрам, а в бассейне Колымы –
лиственничным лесам. Только жалкие следы былой растительности можно сейчас видеть
на южных крутых склонах гор и уступах древних речных террас в разных частях
Чукотки, включая и остров Врангеля.
Таким образом, на протяжении третичного и четвертичного периодов Чукотка как часть
Берингии был колыбелью не только многих видов растений, но и целых
флористических комплексов. Здесь, в Берингии, зародился и сформировался
комплекс листопадных широколиственных лесов (ильмы, дубы, платаны, вязы, клены,
грабы). Листопадность и появилась впервые на Земле как приспособление к
прохладной Берингии. На следующем этапе здесь возникли бореальные виды –
флористический комплекс темнохвойной тайги: все наши и аляскинские ели,
брусника, голубика, багульник и др. Затем идет черед видов-гипоарктов. Наконец,
в последний свой цикл Берингия формирует флористический комплекс тундры. Первое предположение о становлении флоры Арктики
в Берингии высказал в 1937 г. Г. Стеффен. Но окончательное обоснование этой
гипотезы было сделано значительно позднее в работах А.И. Толмачева, Б.А. Юрцева
и других ботаников.
Анализ современных тенденций динамики растительного покрова
северо-Востока в геологическом времени показывает, что сейчас имеет место
типичная для межледниковья и мощной трансгрессии активизация видов и
флористических комплексов, приспособленных к гумидному климату, и регрессия
криоксерофитных видов и сообществ. В будущем можно ожидать расширение ареалов
лугов, каменнобереэовых лесов, появление еловых лесов в Охотии, активизацию
кедрового стланика в континентальных районах, возможно, полную деградацию
степных элементов в растительном покрове как Колымы, так и Чукотки. На Восточной
Чукотке начнут формироваться кустарниковые тундры с доминированием ольховника и
высоких ив, в долинах рек появятся чозения и тополь. Это многовековое развитие
растительного покрова будет, несомненно накладываться на динамику
растительности в экологическом времени, то есть на сукцессии. Сукцессии же,
благодаря масштабным антропогенным нарушениям, будут, в свою очередь, ускорять
это многовековое развитие, снимая так называемое ценотическое торможение
растительного покрова, особенно характерное для него в климаксовом или близком
к климаксовому состоянии.
Литература
1. Атлас Арктики. М.: Изд-во ГУГК, 1985. С. 120–121.
2. Арктическая флористическая область / Под ред. Б.А. Юрцева.
Л.: Наука, 1978. 166 с.
3. Афонина О. М. Листостебельные мхи Чукотского полуострова:
Препр. Магадан: ИБПС ДВО АН СССР, 1988. Ч. 1, 2.
4. Бакаева М. А., Галанин А.В. Об экологической роли
лишайникового покрова в борах-беломошниках средней Вычегды (Коми АССР) //
Экология. 1985. № 2. С. 25–30.
5. Благодатских Л.С. Листостебельные мхи Колымского нагорья:
Препр. Магадан: ИБПС ДВНЦ АН СССР, 1984. 45 с.
6. Васильев В.Н. Растительность Анадырского края. М.; Л.:
Изд-во АН СССР, 1956. 218 с.
7. Геоботаническая карта СССР / Сост. Б. Н. Городковым, Т.
И. Исаченко и др.; Под ред. Е. М. Лавренко и В. Б. Сочавы. М 1: 4000 000. Л.:
Изд-во Ботан. ин-та АН СССР. Ч. 1. 1954. Ч. 2 1956.
8. Егорова Г. Н. Морфолитосистемы и ландшафтная структура
(на примере бассейна р. Омолон). Владивосток: ДВНЦ АН СССР, 1983. 164 с.
9. Колесников Б. П. Растительность // Дальний Восток. М.: изд-во
АН СССР, 1961. С. 183–295.
10. Котляров И. И. Северная материковая часть (Магаданская
область) // Леса Дальнего Востока М.: Лесн. пром-стъ, 1969. С. 199-211.
11. Ниценко А. А. Растительная ассоциация и растительное
сообщество как первичные объекты геоботанического исследования. Л: Наука, 1971.
184 с.
12. Пармузин Ю.П. Северо-Восток и Камчатка. Очерк природы. М.:
Мысль, 1967. 368 с.
13. Петровский В. В. Сосудистые растения острова Врангеля:
Препр. Магадан: ИБПС ДВО АН СССР, 1988. Ч. 1,2.
14. Реутт А. Т. Растительность // Север Дальнего Востока. М.:
Наука, 1970. С. 257–299.
15. Стариков Г. Ф. Леса Магаданской области. Магадан, 1958.
223 с.
16. Стариков Г. Ф., Дьяконов П. Н. Леса Чукотки. Магадан,
1955. 112 с.
17. Хохряков А П. Убежища элементов флоры на севере
Охотского верхнего течения р. Колымы // Бюлл. МОИП, вып. 6. С. 84–96.
18. Хохряков А П. Флора Магаданской области. М.: Наука,
1985. 396 с.
19. Юрцев Б. А Проблемы ботанической географии
Cеверо-Восточной Азии. Л: Наука, 1974. 159 с.
20. Юрцев Б. А Реликтовые степные комплексы Северо-Восточной
Азии. Новосибирск: Наука, 1981. 168 с.
21. Юрцев Б. А, Петровский В. В., Коробков А. А., и др.
Обзор географического распространения сосудистых растений Чукотской тундры.
Сообщ. 1, 2 // Бюлл. МОИП. отд биол. 1979. Т, 84. Вып. 5. С. 11-122; Вып. 6. С.
74–83.