Вы находитесь: НАША БОТАНИЧКА — Растительность — РАСТИТЕЛЬНЫЙ ПОКРОВ СЕВЕРО-ВОСТОКА РОССИИ
РАСТИТЕЛЬНЫЙ ПОКРОВ СЕВЕРО-ВОСТОКА РОССИИ
А.В. Галанин, А.В. Беликович © 1996
Данная работа была написана
в 1996 г., отредактирована проф. С.С. Харкевичем и подготовлена к публикации в составе выпуска 6 Трудов НИЦ "Чукотка". Однако по разным причинам данный выпуск трудов так и не увидел свет. Часть разделов из первой и четвертой глав, сокращенные или дополненные, вошли в главу "Растительный покров" в книге "ПРИРОДА И РЕСУРСЫ ЧУКОТКИ" (1997), некоторые схемы попали в книгу А.В. Беликович "Ландшафтная флористическая неоднородность растительного покрова (на примере модельных районов Северо-Востока России)" (2001). В настоящем электронном ресурсе приводится оригинальный текст целостного очерка, сохранившийся в архиве авторов.
Пояснение терминов
Чтобы читатель мог прочесть именно то, что хотели сказать
авторы, следует привести словарь специальных терминов и понятий. Под Северо-Востоком России в данной работе понимается регион, включающий в себя два административных подразделения РФ – Магаданскую область и Чукотский автономный округ.
Растительный покров – это множество особей растений, которые
мы относим к разным видам, родам, семействам, порядкам, классам и типам
(таксономическая классификация) и разным жизненным формам, экологическим
группам, сообществам (экологическая классификация). Растительный покров неоднороден,
он изменяется в пространстве – от места к месту, и во времени – от года к году.
При этом изменяется его видовой состав – флора – и набор жизненных форм,
экологических групп и сообществ – растительность.
Растительное сообщество (фитоценоз) это относительно
однородный участок растительного покрова ранга физико-географической фации
(порядка нескольких сотен квадратных метров). На этом участке
микроклиматические, гидрологические и почвенныe условия варьируют
незначительно, здесь на всем протяжении выдержан один набор видов и жизненных
форм растений, которые, определенным образом сочетаясь друг с другом, образуют
относительно устойчивые группировки – ценоэлементы – так, что одно растительное
сообщество может состоять из одного, двух, иногда трех и более типов ценоэлементов,
закономерно сочетающихся в пределах контура этого сообщества.
Характеризуя растительность нашего региона, мы будем
использовать два типа ценоэлементов – ценоячейки и синузии. Синузия – это слой,
или чаще всего ярус растительного покрова, состоящий из особей растений сходной
жизненной формы, например: синузия кедрового стланика, синузия сфагновых мхов,
синузия гипоарктических кустарничков (голубика + багульник + мирт болотный +
брусника + шикша). Ценоячейка – это элемент микромозаики растительного покрова
размером в несколько квадратных метров, образованный особью или группой особей доминантов
и сопутствующих им подчиненных особей. Особь-лидер в ценоячейке влияет на
среду, а через нее и на окружающих соседей, которые зачастую настолько зависят
от лидера, что вне его фитогенного паля "чувствуют" себя не вполне
уютно. Именно набор ценоячеек и синузий определяет характер растительного
покрова как компонента экосистем и биосферы. Нередко особи разных видов мы
относим к одной синузии – разные виды сфагновых мхов на болоте; разные виды
разнотравья в пойменном чозениевом лесу. Напротив, особи одного вида могут образовывать
разные синузии, как например, синузия высокорослых кустарников кедрового
стланика (стланиковые леса) в нижних частях горных склонов и синузия
низкорослых тундровых кустарников из того же стланика в привершинных частях
склонов. Жизненная форма – это не просто внешний облик растения или форма его
роста, это его экологическая и ценотическая стратегия в сообществе и экосистема.
Растительная aссоциация, или эколого-ценотический элемент
локальной флоры – это набор видов растений, которые в растительном покрове ландшафтного
района чаще встречаются вместе друг с другом в одних фитоценозах и реже порознь
в разных фитоценозах. Растительную ассоциацию часто рассматривают как тип
фитоценозов, хотя с логической точки зрения это не совсем правильно.
Неоднородность растительного покрова в пространстве
прослеживается на многих уровнях. Обычно рассматривают уровень фитоценозов и микрокомбинаций
(фации), уровень мезокомбинаций (урочища), мaкpoкомбинаций (местности или
элементарные геоботанические районы), мегакомбинаций (геоботанические провинции
и области). Карты растительного покрова (геоботанические карты), как правило,
отражают неоднородность растительного покрова на уровне микро- и
мезокомбинаций, а схемы геоботанического районирования – на уровне макро- и мегакомбинаций.
Виды растений в растительном покрове представлены множествами особей разного
ранга, или популяциями: ценопопуляция – все особи одного вида в пределах
фитоценоза; местная популяция – все особи одного вида в пределах
геоботанического района или геоботанической провинции. Особи в одной популяции
генетически связаны друг с другом, они могут скрещиваться и имеют единый
генофонд популяции.
Заселяя какое-то местообитание, свободное от растительности,
особи растений вступают друг с другом в конкурентные и всевозможные
неконкурентные, в том числе и аллелопатические отношения. В результате этих
взаимодействий в растительном покрове реализуются далеко не все статистически
возможные сочетания видов данной локальной флоры некоторые такие сочетания
элиминируются как ценотически и экологически неоптимальные. Постепенно
формируется устойчивое растительное сообщество с хорошо выработанной ценотической
структурой, оптимально выполняющее в экосистеме свою биогеохимическую функцию.
Пионеpное сообщество сменяется серийным cooбщecтвом, а то, в свою очередь,
климаксовым coo6щecтвом. Так происходит растительная сукцессия (смена), в ходе
которой в растительном покрове вырабатывается все более тонкая ценотическая
структура. Сукцессионные процессы мы наблюдаем обычно там, где климаксовое
сообщество в той или иной мере было разрушено или выведено из равновесия
человеком или каким-то природным фактором.
Введение
Северо-Восток России в ботаническом отношении – регион необыкновенный,
так как он входит в состав Берингийского сектора Арктики и Субарктики. Бурная
геологическая история Берингии наложила свой отпечаток на растительный покров
региона, сделав его колыбелью многих видов и целых ассоциаций растений, ныне
широко расселившихся в тундровой и таежной зонах 3емли.
В сводке А.П. Хохрякова "Флора Магаданской
области" [18] для Магаданской области с Чукоткой приводится 1500 видов
сосудистых растений. Видовое разнообразие растительного покрова только Чукотки,
по данным Б.А. Юрцева с соавторами [21], свыше 900 видов. Даже флора острова
Врангеля, по данным В.В. Петровского [13], насчитывает не менее 385 видов, что
значительно больше флоры любого равновеликого острова в Арктическом океана
Флора мхов магаданской области и Чукотки изучена в меньшей
степени, чем флора сосудистых растений. Однако приблизительное число видов на
основании исследований О. В. Афониной и Л.С. Благодатских можно оценить цифрой
не менее 600 для всего региона и около 400 – только для Чукотки. Флора
лишайников в нашем регионе изучена еще более фрагментарно. Но, основываясь на
данных И.А. Макаровой и Ю.А. Котлова, общее видовое разнообразие лишайников
магаданской области и Чукотки можно оценить примерно в 400–500 видов.
Таксономически или, точнее, флористически территория
северо-Востока России весьма неоднородна. Существует несколько вариантов ее
флористического районирования. Среди них наиболее известны: районирование,
принятое в многотомной сводке "Флора СССР"; районирование во
"Флоре магаданской области" А.П. Хохрякова [18]; районирование Б.А.
Юрцева [19]; и районирование, принятое С.С. Харкевичем в многотомной сводке
"Сосудистые растения советского Дальнего Востока".
Дифференциация растительного покрова региона по
растительности, естественно, отличается от его дифференциации по флоре. К
сожалению, хорошо разработанных и обоснованных схем геоботанического
районирования, и тем более геоботанических карт удобных для пользователя
масштабов на наш регион пока нет. Наиболее современны мелкомасштабные карты,
входящие в Атлас карт растительности СССР под редакцией Е.М. Лавренко и В.Б.
Сочавы [7], а на Арктическую область – карта, выполненная С.А. Грибовой в
"Атласе Арктики СССР" [1].
Если же говорить о крупных масштабах, то имеются карты
растительности лишь отдельных ключевых участков А.Е. Катенина, С.С. Холода,
А.В. Беликович, Ю.Б. Королева и др. Имеются также мелкомасштабные геоботанические
схемы распределения растительности, такие например, как карта А.Т. Реутт [14]
или В.Н. Васильева [6]. Для характеристики растительности можно успешно
пользоваться ландшафтными картами и схемами ландшафтного районирования Г.Н.
Егоровой [8].
На основании личных исследований и опубликованных материалов
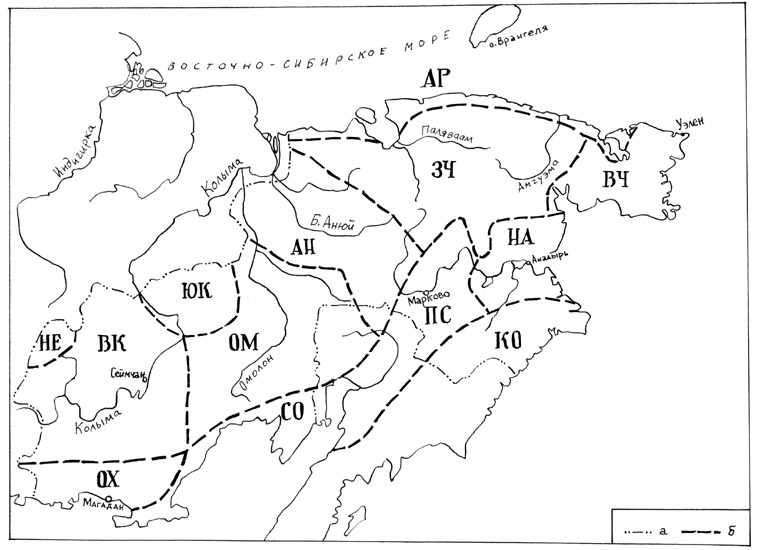
мы предлагаем схему геоботанического районирования Северо-Востока России (рис.
1). В основу нашего районирования в первую очередь положена идея
широтно-долготной дифференциации растительного покрова под воздействием климата
и его дифференциация в связи с различиями в литогенной основе и рельефе (мега-
и макрорельеф). При проведении геоботанических границ (границ геоботанических
округов) мы не учитывали градиенты флористического районирования, за
исключением, пожалуй, границ распространения видов-доминантов и эдификаторов
(то есть мощных ценообразователей – видов, особи которых в процессе
жизнедеятельности заметно влияют на среду обитания и трансформируют ее в новое
экологическое состояние).
|
На основе широтной и долготной дифференциации растительного
покрова нами выделена зона арктических тундр, куда входит Арктический округ;
зона типичных и южных гипоарктических тундр и лесотундры с округами Западно-Чукотским,
Восточно-Чукотским, Нижнеанадырским тундровым и Пенжинско-Среднеанадырским,
Корякским и Ceвepo-Охотским лесотундровыми; зона лиственничной тайги с округами
Анюйским, Омолонским, Юкагирским, Верхнеколымским, Нерским и Охотским С востока
на запад геоботанические округа образуют своеобразные ряды усиления
океаничности. Первый ряд образуют Охотский, Верхнеколымский и Нерский геоботанические
округа; второй ряд – Северо-Охотский, Омолонский и Юкагирский; третий ряд –
Нижнеанадырский, Пенжинско-Среднеанадырский и Анюйский; четвертый ряд
Восточно-Чукотский и Западно-Чукотский геоботанические округа. Корякский округ,
в силу своего географического положения, выпадает из этой схемы долготной
геоботанической зональности; он скорее является продолжением полуострова
Камчатка.
Слева: Рис. 1. Геоботаническое районирование Северо-Востока России (авторская схема). а – граница Северо-Востока России; б – границы геоботанических округов.
Геоботанические округа: АР – Арктический, ЗЧ – Западно-Чукотский, ВЧ – Восточно-Чукотский, НЕ – Нерский, ВК – Верхнеколымский, ЮК – Юкагирский, ОМ – Омолонский, АН – Анюйский, ОХ – Охотский, СО – Северо-Охотский, ПС – Пенджинско-Среднеанадырский, НА – Нижнеанадырский, КО – Корякский |
История
ботанических исследований
Самые первые ботанические исследования в регионе связаны с
Великими географическими экспедициями на Северо-Востоке Азии – Северной
экспедицией А.И. Чирикова (1732–1743 гг.), экспедицией И.И. Биллингса и Г.А.
Сарычева (1785–1793 гг., первые коллекции Мерка), экспедицией Ф.П. Врангеля
(1820–1824 гг.), Чукотской экспедицией 1868–1870 гг. Большой вклад в пополнение
данных о растительности в XIX в. внесли путешественники, географы и геологи,
отдельные натуралисты, священники-миссионеры, представители русской военной и
гражданской службы. Благодаря им в разных гербариях, музеях и ботанических
садах страны и мира стали формироваться коллекции видов растений, собранных на
Северо-Востоке Азии. Так, приближенное суждение о флоре Верхней Колымы давали
растения, собранные врачом Ф. Августиновичем в 1875–1876 гг. и обработанные
Е.Р. Траутфеттером; о флоре Анадырской округи – гербарий, собранный начальником
этой округи Л.Ф. Гриневецким в 1890 г. и позднее обработанный известным
ботаником В.Б. Сочавой.
Серьезное и планомерное изучение растительного покрова
Северо-Востока началось в 1930-е годы в связи с заказами Акционерного
Камчатского общества, Дальстроя, Народного комиссариата земледелия РСФСР,
особенно в связи с изысканиями Государственного землеустроительного треста.
Среди первых геоботанических работ следует назвать исследования В.Б. Сочавы в
бассейнах р. Анадырь и р. Белая, В.Н. Васильева в бассейне р. Анадырь, Л.Н.
Тюлиной в верхнем течении р. Анадырь, р. Майн, Б.Н. Городкова в Пенжинском
районе, по берегам бухты Провидения, залива Лаврентия, в районе мысов Ванкарем
и Шмидта и на острове Врангеля.
Широко развернувшиеся в конце 1930-х годов работы по
землеустройству территории, организации лесоучетных работ, изучению кормовых
ресурсов оленьих пастбищ интенсивно продолжались до начала 1960-х годов. Эти
работы включали обязательное обследование растительности, составление
геоботанических карт, аэротаксацию. Из наиболее крупных геоботанических работ,
вышедших в эти годы, можно привести интересные фенологические наблюдения А. П.
Васьковского в Примагаданье и его обзор ареалов основных ценозообразователей
магаданской области, описание лесов региона С. К. Старка, а позднее – Б. П.
Колесникова, Г. Ф. Старикова и П. Н. Дьяконова [10, 11], работы Б. А Тихомирова
по кедровому стланику и лесотундрам.
После окончания эпохи сталинских репрессий и постепенного
закрытия и переустройства лагерей регион стал интенсивно посещаться
академическими экспедициями. Начиная с конца 1960-х годов, большую работу по
изучению флоры и растительности Чукотки провели сотрудники лаборатории растительности
Крайнего Севера Ботанического института АН СССР им. В.Л. Комарова, сначала под
руководством Б.А. Тихомирова, а затем Б.А. Юрцева. Значительная часть
материалов, собранных ими в многочисленных экспедициях на протяжении 20 лет,
опубликована в многотомной сводке "Арктическая флора СССР", в
нескольких монографиях и многочисленных научных статьях (Б. А. Тихомиров, В. А
Гаврилюк, Б. А. Юрцев, В. В. Петровский, А. А. Коробков, Ю. П. Кожевников, Т. Г.
Полозова, Т. М. Заславская, В. Ю. Разживин и др.). На юге региона планомерную
инвентаризацию флоры провел коллектив лаборатории ботаники Института
биологических проблем Севера под руководством А.П. Хохрякова (Т. М. Мазуренко, Л.
С. Благодатских, А.Н. Беркутенко, Г. Л. Антропова и др.). Однако вплоть до
начала 1980-х годов доминировали флористические исследования, собственно
геоботаникой занимались немногие ученые (Ю. П. Кожевников, Т. Г. Дервиз-Соколова,
А. К Катенин). Хорошие обзоры растительности региона можно найти в сводках
"Дальний Восток" [9], "Северо-Восток и камчатка" [12],
"Леса Дальнего Востока" [10], "Север Дальнего Востока" [14].
С середины 1980-х гг. детальным изучением структуры
растительного покрова на Чукотке стала заниматься лаборатория ботаники в
Магаданском институте биологических проблем Севера ДВО РАН, а позднее – лаборатория
структуры растительного покрова того же института, делающая акцент на изучение
принципов организации растительного покрова, его роли в биологическом
круговороте и проводящая разнообразные ресурсные исследования. Настоящая сводка
написана в духе концепции растительного покрова, разработанной в этой
лаборатории.
Основные синузии (экологический аспект растительного
покрова)
Растительность Северо-Востока России представлена немногими,
но ясно выделяющимися типами сообществ. Для каждого типа растительности
характерны свои доминантные виды, ценоячейки и синузии, своя стратегия биологического
круговорота вещества. Разнообразие растительности в ландшафте весьма тесно
скоррелировано с характером рельефа, гидрологическим режимом, микроклиматом,
минералогическим составом горных пород и почвами, микробным, грибным и животным
населением. В совокупности с этими компонентами и факторами растительность
образует локальные экосистемы, или биогеоценозы. Для того чтобы охарактеризовать
типы растительности, а сделать это можно только характеризуя синузиальное
сложение сообществ, мы должны детально остановиться на описании основных синузий.
Синузии мхов. Все синузии мхов
– это тонкий, но весьма плотный ковер споровых растений, однако все они имеют
свои отличия.
Синузия сфагновых
мхов – это очень высокогигроскопичный ковер толщиной до 20–30 см. Наиболее часто в составе синузии сфагновых мхов на
Северо-Востоке России встречаются следующие виды сфагнов: Sphagnum fuscum, Sphagnum lenensе, Sphagnum magellanicum, Sphagnum lindbergii, Sphagnum warnstorfii, Sphagnum squarrosum,
Sphagnum compactum, Sphagnum teres, Sphagnum girgensohnii, Sphagnum russsowii, Sphagnum fimbriatum [5]. Всего в регионе
встречается более 30 видов сфагновых мхов. Видовой состав сфагновой синузии
изменяется в зависимости от интенсивности и постоянства переувлажнения, а также
от степени минерализованности воды, питающей сфагновую толщу.
Синузия зеленых
мхов – это также тонкий и плотный ковер
растений, играющий большую роль в экосистемах Арктики и Субарктики. Имея
большой запас фитомассы на единицу площади, зеленые мхи влияют на процесс
движения вещества и превращения энергии в экосистемах. Они выполняют здесь
стабилизирующую, буферную роль, обеспечивая экологическое и биогеохимическое
равновесие, формируя специфическую среду обитания для всех других растений,
микроорганизмов, грибов и животных.
Таксономически и экологически синузия зеленых мхов весьма
неоднородна. Вполне вероятно, что она может разделяться специалистами-бриологами
на несколько синузий в зависимости от плотности и гигроскопичности мохового
ковра, темпов прироста и разложения бриомассы. В разных районах региона и на
разных местообитаниях в состав синузии зеленых мхов могут входить следующие
виды: Роlytriсhum commune, Роlytriсhum alpestre, Роlytriсhum jensenii, Роlytriсhum piloferum, Роlytriсhum juniperinum,
Drepanocladus uncinatus, Drepanocladus fluitans, Drepanocladus ехаnnulatus, Racomitrium саnescens, Racomitrium lanuginosum, Pleurozium schreberi, Dicranum polysetum, Dicranum majuus, Dicranum affine, Dicranum еlongаtum, Dicranum congestum, Aulacomnium palustre, Aulacomnium turgidum, Hylocomium splendens, Brachytecium spp., и др. [16]. Всего бриофлора
северо-Востока России, по данным Л. С. Благодатских и О. М. Афониной,
насчитывает более 500 видов и рас листостебельных зеленых мхов. Только на
Чукотском полуострове обитает 359 видов листостебельных мхов (включая
сфагновые, гипновые и бриевые мхи [3]).
Синузия гипновых
мхов во многом подобна синузиям cфaгновых и
зеленых мхов. Это также плотный и тонкий ковер миниатюрных растений, встречающихся
во многих экосистемах региона. Таксономическое разнообразие синузии гипновых
мхов не меньше, чем зеленых. Это Leptobryum pyriforme, Мniobryum wahlenbergii, разные виды родов Вryum, Мnium,
Нypnum, Calliergon и др. По всей вероятности, следует выделить отдельную синузию
водных мхов, образующих обрастания камней на дне рек и ручьев. В основном это
разнообразные представители родов Нydrohypnum. Наличие
в экосистеме той или иной из четырех рассмотренных нами моховых синузий четко
индицирует особенности биологического круговорота и во многом определяет
условия произрастания других растений из других синузий.
Синузии лишайников подобно моховым, представляют собой плотный и тонкий ковер споровых
растений, но совершенно своеобразных по своей биологии. Это так называемые
организмы-симбионты, в теле которых сожительствуют два разных существа –
многоклеточный гриб и одноклеточная водоросль. Нити, или гифы гриба, оплетают
клетки водоросли, которые на свету из воды, углекислого газа и небольшого
количества минеральных элементов синтезируют органическое вещество. Сам гриб не
фотосинтезирует, это делает водоросль, но гриб всасывает из внешней среды воду
с растворенными в ней веществами и по своим гифам предает ее клеткам водоросли.
Грибные гифы в сырую погоду вбирают в себя много воды, которую затем медленно
расходуют и отчасти передают ее водоросли.
Лишайники
устойчивы к периодическому высыханию. Они не менее гигроскопичны, чем сфагновые
мхи, и во время дождей способны запасать много воды, которую затем постепенно
расходуют в сухую погоду. Именно эта особенность позволяет им занимать сухие местообитания
– часто каменистые и щебнистые. С помощью органических лишайниковых кислот
грибные гифы лишайника способны растворять горные породы и минералы.
Синузия кустистых
и листоватых лишайников зачастую
доминирует во многих экосистемах континентальных горных районов как лесной, так
и тундровой зон. Нами установлено, что лишайниковая синузия в
северных лесах оптимизирует биологический круговорот, ускоряя его и делая боле
замкнутым [4]. На севере сплошной ковер лишайников встречается во многих
экосистемах. Обладая низкими темпами прироста (несколько миллиметров в год),
лишайники, тем не менее, накапливают значительную биомассу, так как отмирая,
разлагаются очень медленно. В отличие от мхов, синузия лишайников играет
большую роль в трофических цепях. Кустистые лишайники – важная часть рациона
дикого и домашнего северного оленя, некоторых видов мышевидных грызунов.
Ресурсы зимних оленьих пастбищ в регионе значительны. В
некоторых районах, как например, на Чукотском полуострове и в восточной части
Анадырского административного района, из-за перевыпаса эти ресурсы сильно
подорваны, что катастрофически отразилось на оленеводстве численность оленей
здесь за последние 10–15 лет сократилась более чем в 4 раза. Чтобы восстановить
деградировавшие лишайниковые пастбища, природе потребуются многие десятилетия,
хотя для разрушения лишайниковой синузии путем перевыпаса достаточно одного года,
а то и просто одного месяца.
Наиболее характерными видами, слагающими у нас эту синузию,
являются Cladonia alpestris, Cladonia rangiferina, Cladonia sylvatica, Cetraria nivalis, Cetraria cucullata, Cetraria islandica, Cetraria laevigata, Alectoria оchroleuca, Alectoria nigricans, Peltigera aphtosa, Peltigera ochroleuca, и др. Всего в составе
этой синузии на северо-Востоке России насчитывается не менее 60–80 видов.
Синузия накипных,
чешуйчатых и корковых лишайников представляет собой тонкую, плотно приросшую
пленку на поверхности скал, камней, щебенки, уплотненной почвы, органических
остатков и других субстратах. Огромные пространства каменных россыпей на вершинах
и склонах гор на Чукотке заняты этой синузией. Накипные лишайники очень
медленно растут, возраст одной особи – овального серого пятна лишайника рода Lecidea диаметром 40 см на поверхности скалы может быть более 2000 лет, а особи
накипного зеленоватого лишайника вида ризокарпон географикум диаметром в 15 см
– значительно превышает 1000 лет. Таксономический состав синузии накипных
лишайников изменяется в зависимости от химизма субстрата, характера горных
пород, высоты над уровнем моря и зонального положения района. Всего в регионе в
составе этой синузии встречается более 300 видов лишайников. Наиболее обычны
представители родов Rhizocarpon, Parmelia, Lecidia,
Parpidia и др.
По всей вероятности, накипные, чешуйчатые и корковые
лишайники образуют не одну, а две или даже три синузии, достаточно разные по
экологической значимости, однако здесь мы ограничимся, пожалуй, упоминанием
лишь одной эпифитно-лишайниковой синузии на стволах и сучьях живых деревьев и
кустарников. Эта синузия очень важна в обеспечении биологического круговорота,
так как лишайники-эпифиты (чаще всего из рода Parmelia) перехватывают подвижные
химические элементы, освобождающиеся при разрушении коры деревянистых растений,
не давая этим элементам попасть в почву и почвенные воды. Интересно, что в приморской зоне региона, где выпадает
много осадков в виде дождя и угроза вымывания подвижных химических элементов из
почвы очень велика, роль лишайников-эпифитов в экосистеме резко возрастает, и в
лесу мы всюду встречаем космы бородача (Usnea), свисающие с сучьев и ветвей
деревьев.
Синузии трав распространены на Северо-Востоке России гораздо менее широко, чем мохово-лишайниковые. Они делятся на несколько синузий в зависимости от отношения к влаге и ее солености.
Синузия
мезофильных трав на Северо-Востоке России
распространена не очень широко, хотя с ней мы постоянно сталкиваемся в долинах
рек, на южных склонах гор. В некоторых экосистемах синузия мезофильных трав не
сочетается с другими синузиями – это настоящие луга; в других она идет под
пологом тополя и чозении, разных видов ив, каменной березы, то есть образует
вместе с ними сложные многоярусные и полисинузиальные растительные сообщества. Сюда входит очень много видов травянистых
растений, чаще всего многолетних коротко- и длиннокорневищных, дерновинных,
розеточных, реже луковичных. Вся наземная часть этих растений
ежегодно отмирает и ежегодно возобновляется. Мезофильными считаются травы, которые хорошо растут при
нормальном увлажнении на достаточно плодородных почвах в местах, не очень
сильно промерзающих зимой.
Синузия мезофильных трав обеспечивает в экосистемах
интенсивный биологический круговорот, так как мертвые остатки трав быстро
разлагаются, а зеленая масса, как правило, хорошо поедается животными, что
также ускоряет оборот вещества в экосистеме. Даже в течение одного
вегетационного периода химические элементы, попадающие с травой в кишечный
тракт животных на лугу, могут пройти через круговорот дважды или тpижды. Ежегодная
продукция вещества травянистой синузией в магаданской области может составлять
до 40–50 ц/га воздушно-сухого веса в пойменных лугах и до 15–20 ц/га
воздушно-сухого веса в лесах с развитым травяным покровом.
Нивальные луговины в зоне тундры на Чукотке могут ежегодно
производить до 10–15 ц/га воздушно-сухого вещества, что на 2–3 порядка больше,
чем в других типах тундровой растительности. Синузия мезофильных трав в регионе
включает много видов травянистых растений и заметно изменяется от местообитания
к местообитанию, от зоны к зона Известны крупнотравные разнотравные и злаковые,
плотнодерновинные и рыхлодерновинные, редкотравные (на галечниках рек) и другие
варианты этой синузии. Очень часто в составе этой синуэии преобладает какой-то
один вид (вейник Лангсдорфа, овсяница алтайская, костер Пампэлла и др.) или
несколько видов образуют смешанное население синуэии (например, купырь лесной,
волжанка камчатская, дудник и другие травы в каменноберезовом лесу). Всего в
этой синузии нами зарегистрировано свыше 500 видов.
Сообщества, в которых развита синузия травянистых мезофитов,
имеют большое практическое значение. Это пастбища для крупного рогатого скота и
оленей, после расчистки сенокосные луга, места заготовок ряда лекарственных
растений. Кроме того, все экосистемы, где обильно растут травянистые мезофиты,
отличаются интенсивным биологическим круговоротом и имеют высокий экологический
потенциал. Здесь лучшие почвы, благоприятный микроклимат, нормальное увлажнение
и др.
Синузия гидро- и
гигрофильных трав существенно отличается от
предыдущей. Ее образуют травянистые растения, имеющие экологический оптимум в
постоянно переувлажненных местообитаниях – мелководьях озер, заводях, старицах
рек, низких затапливаемых берегах водоемов и отмелях. Внешне эта синузия бывает
похожа на синузию травянистых мезофитов, и в этом случае ее нередко называют заболоченным
лугом. Однако, в отличие от лугов, в таких местах продукция органического
вещества превышает разложение и идет накопление торфа. С одной стороны, это связано
с частым спутником – синузией гипновых мхов; мхи определяют процесс
торфонакопления, а травы вовлекаются в него. С другой стороны, физиология и
вообще биология водолюбивых травянистых растений очень отличается от таковой
травянистых мезофитов. Например, в корневищах эти растения, как правило, имеют
воздухоносную паренхиму, плоды и семена у них чаще всего распространяются
водными потоками, имея к этому целый ряд приспособлений. Многие из
прибрежно-водных трав накапливают физиологически-активные вещества,
воздействующие на травоядных животных.
Пожалуй, в одном гидро- и гигрофиты сходны с мезофитами – и
те, и другие образуют большую ежегодно отмирающую надземную фитомассу, хорошо
поедаются травоядными животными и обеспечивают интенсивный круговорот химических
элементов. Среди синузии гидро- и гигрофильных трав также много видов из разных
родов и семейств, но в целом ее таксономическое разнообразие несколько меньше,
чем синузии мезофитов. Наиболее обычны здесь виды рдестов (Роtamogeton), осок (Carех), хвощей (Equisetum palustre,
Equisetum limosum), водяная сосенка (Нippuris), водяная звездочка (Callitriche), сабельник болотный (Сomarum palustre), калужница (Caltha), и др. Здесь встречается много лекарственных
растений, некоторые из них могут заготавливаться в промышленных масштабах.
Синузия ксерофильных
и петрофильных трав встречается довольно редко. Это так называемая степная, степоидная и лугостепная
растительность в резко континентальных районах Колымы и Чукотки, а также
травянистый покров в разреженных лиственничниках на крутых южных склонах гор в
тех же районах. В состав этой синузии входит ряд видов, ареалы которых
простираются в зону степей Азии и Европы. Это ксерофильные злаки, осоки,
разнотравье. Однако наряду со степными видами здесь встречаются и довольно
обильны виды-петрофиты – камнеломка (Saxifraga funstonii),
тимьян (Тhymus serpyllum s.l.), змееголовник (Dracocephalum palmatum); мезо-ксерофиты –
подмаренник (Galium vеrum), горец (Polygonum laxmannii), иван-чай узколистный (Chamaerion
angustifolium), и др.
То, что в Магаданской области синузия ксерофильных трав образует реликтовый тип
растительности, считают многие ботаники [19]. Однако безоговорочное отнесение
этих сообществ к степям по ряду соображений весьма проблематично. Например, в окрестностях
г. Сусуман на так называемых степных склонах мы постоянно наблюдаем следы
лесных пожаров обгоревшие стволы и пни лиственницы и кедрового стланика. Очень
похоже, что некоторые южные склоны, покрытые ксерофильной травянистой растительностью
это стадия замедленной послепожарной сукцессии. Подтверждение этой гипотезе
можно найти в исследованиях почвоведов, утверждающих, что так называемые кутаны
(отложения солей на поверхности щебенки в почве, возникающие в результате
испарения почвенных растворов и всевозможных химических взаимодействий этих
растворов с минералами горной породы) имеют здесь своеобразную слоистость.
Kapбонатные прослойки, формировавшиеся под степоидной
растительностью, чередуются с силикатными прослойками, формировавшимися под
лесной растительностью (устное сообщение Е.М. Наумова). Длительность этого
сукцессионного цикла – около 250–300 лет. Таким образом, фактором,
поддерживающим постоянное наличие в континентальных ландшафтах синузии
травянистых ксерофитов как сукцессиннной стадии, является периодическое выгорание
лиственничных лесов.
Синузия
плотнодерновинных трав-кочкообразователей характерна для местообитаний с хотя бы периодическим застойным увлажнением
в зонах тундры и тайги. Высокие кочки (высотой иногда до 1 м) могут занимать
огромные площади, в эначительной степени определяя облик ландшафта и условия
жизни других растений. Кочкообразователями в нашем регионе могут быть некоторые
визы осок (Сarех cespitosa, Carex wiluica, Carex appendiculata, Carex
lugens, Carex arctisibiriса, Carex soczavaeana, и др. ), пушица влагалищная
(Еriорhorum vaginаtum), овсяница алтайская (Festuca altaica)
и некоторые другие плотнодерновинные травянистые растения.
Наряду с кочкарными пушицевыми тундрами и кочкарными
осоково-кустарничковыми лиственничными редколесьями (марями) в регионе часто
встречаются так называемые кочкарные осоковые луга в долинах рек и ручьев в
лесной зоне. Кочкообразование здесь также связано с особой жизненной формой
растений, поселяющихся в переувлажненных местах. Наряду с оптимизацией
продукционного процесса кочкообразование здесь, по-видимому, выполняет и
функцию биогеохимического фильтра: во время паводка такая растительность
способствует осаждению взвесей и коллоидов.
Формированию кочек в переувлажненных местах способствует
перевыпас коров и оленей. В таких случаях этот процесс также можно
рассматривать как защитным механизм экосистемы, срабатывающий в экстремальных
условиях и направленный на сохранение в экосистеме биогеохимического и
продукционного потенциала. Все
пространство такого луга оказывается разделенным на кочки, куда животные не
ступают (это просто неудобно) и межкочечные пространства, которые и утаптываются
животными. Растения в этом случае растут в кочках и на кочках, и животными
угнетаются значительно меньше.
Синузия
травянистых приморских галофитов хорошо знакома тем, кто бывал на низких сырых и умеренно влажных берегах морей. Это плотный упругий
ковер из низкорослых длинно- и короткокорневищных и дерновинных злаков, осок,
ситников и разнотравья в зоне интенсивного засоления. Такая растительность
обычно называется маршевыми лугами (иногда просто маршами). Марши интенсивно
удобряются птичьими экскрементами (это часто гусиные пастбища), значительная
часть растений выедается птицами, поэтому здешние экосистемы относятся к числу
наиболее продуктивных и биогеохимически очень интенсивных. Вместе с птичьим
пометом на маршевые луга выносится значительное количество вещества из моря,
где птицы кормятся рыбой.
Так как синузия травянистых галофитов не сочетается ни с
какими другими синузиями, ее таксономический спектр представляет собой
ассоциацию и обсуждается в разделе об ассоциациях. Набор видов и экобиоморф в
пределах синузии изменяется в связи с изменением интенсивности увлажнения и
степенью засоления.
Синузии кустарничков состоят из
невысоких (до 1 м высотой) многолетних растений с одревесневающими надземными
побегами.
Синузия
гипоарктических кустарничков включает в себя багульники (Ledum palustre, Ledum decumbens) , голубика
(Vaccinium uliginоsum), болотный мирт (Chamaedaphne calyculata), подбел (Andromeda polifolia), брусника (Vaccinium vitis-idaea), шикша (Empetrum nigrum s.l.), карликовая береза (Вetula nаnа subsp.
exilis) и некоторые другие виды. Растения этой синузии отличаются низкой
продуктивностью, поэтому для экосистем с этой синузией характерен замедленный
биологический круговорот. Это типичные олиготрофные экосистемы равнинных тундр,
сфагновых и гипновых болот, лиственничных редин и редколесий. Синузия
гипоарктических кустарничков занимает узловое место в растительном покрове всех
геоботанических округов Северо-Востока России. По этой синузии в северном
полушарии Б.А. Юрцевым [2] выделяется гипоарктический ботанико-географический
пояс – зона, в которой гипоарктические кустарнички играют ведущую роль в так
называемых плакорных местообитаниях, то есть преобладают в ландшафтах.
Считается, что гипоарктические кустарнички особенно активизировались в
ледниковые эпохи в континентальных и резкоконтинентальных районах.
Синузия дриады (Dryas punctata s.l.) встречается в зональных
и горных тундрах. Дриада, или куропаточья трава – это низкорослый распластанный
по земле кустарничек, часто имеющий подушкообразную форму роста, ксерофит с медленным ростом, с
вечнозелеными листьями, функционирующими до 10 лет. В природе
синузия дриады формирует своеобразную пятнистую тундру, в которой между ее
отдельными особями значительная часть щебнистого субстрата остается
незаселенной сосудистыми растениями. На северо-Востоке
России систематики обычно различают несколько видов дриады, близких по своей
жизненной форме, но образующих, на наш взгляд, одну синузию. Это Dryas
octopetala, Dryas punctata, Dryas incisa,
Dryas integrifolia, Dryas grandis, и другие, менее распространенные.
Синузия
кустарничковых ив очень близка и по внешнему облику,
и по структуре и темпам ежегодного прироста к синузии гипоарктических
кустарничков. Однако от типичных гипоарктов кустарничковые ивы отличаются более
высокой ежегодной продуктивностью. Большинство таких ив – не олиготрофы, как гипоарктические
кустарнички, а мезо- и даже эвтрофы. Это ивы клинолистная (Salix sphenophylla),
арктическая (Salix arctica), сетчатая (Salix reticulata), полярная (Salix polaris),
чукотская (Salix tschuktschorum), красивая (Salix pulchra), скальная (Salix saxatilis) и другие.
Ежегодная продукция листьев и сережек в синузии
кустарничковых ив в ряде случаев приближается к ежегодной продукции наземных
частей трав в синузии травянистых мезофитов. Именно поэтому в составе нивальных
лужаек, пойменных и иных лугов в тундровой и лесотундровой зонах мы встречаем
много кустарничковых ив. Хотя в ряде случаев (ивы красивая и клинолистная в
тундрах Чукотки) эти ивы ведут себя в биологическом круговороте скоррее как
олиготрофные гипоарктические кустарнички.
Синузии кустарников.
Синузия березки
Миддендорфа представляет собой заросли высотой до 2 м. Ерники из березки Миддендорфа (именно ерниками называют в
народе эту синузию) очень широко встречаются в лесной и лесотундровой зонах
региона, всегда в местах с неглубоким
залеганием многолетней мерзлоты. В местах, где хорошо развиты синузии
лиственницы или кедрового стланика, заросли березки становятся разреженными или
совсем исчезают. Способность березки разрастаться после
пожаров приводит к тому, что ее синузия начинает преобладать на лесных вырубках в лиственничных лесах и
редколесьях, а также после пожаров. За счет мощной
корневой системы материнского растения, наземная часть которого погибла при
пожаре, корневая поросль быстро опережает в росте все другие деревья и
кустарники.
В отличие от синузии кедрового стланика, ерники образуют менее
олиготрофные экосистемы, так как интенсивность биологического круговорота,
проходящего через березку, значительно вьппе. Продукция однолетней фитомассы
листьев значительна, к тому же и разлагаются эти листья довольно быстро.
Синузия ольхи
кустарниковой (Alnus fruticosa, Alnus kamtschatica, Alnus hirsuta) встречается в разных
геоботанических округах региона, в тундровой зоне становится редкой. Ольха – эвтрофное
(высокопродуктивное) растение, и поэтому селится там, где интенсивность биологического
круговорота повышена (ложбины стока, окраины ключевых болот, места выхода
на поверхность грунтовых вод, обочины дорог, старые зарастающие дороги и
минерализованные полосы – но всегда в местах с достаточным или слегка
избыточным увлажнением). Кустарниковая ольха быстро растет и ежегодно производит большую
фитомассу листьев, опадающих осенью. Заросли ольховника в
благоприятных местах могут достигать 3–4 м высоты и быть очень густыми.
Особи ольховника недолговечны, и в большинстве случаев в сукцессионном ряду на
смену ольховникам приходят сообщества с доминированием лиственницы или
кедрового стланика, в которых синузия кустарниковой ольхи играет подчиненную
роль или исчезает совсем.
Синузия кедрового стланика (Рinus
pumila) в регионе в значительной степени определяет лицо ландшафта. Это густые
заросли стелющегося темнохвойного кустарника высотой до 3,0–3,5 м, типичного
олиготрофа. Хвоя на ветках сохраняется 5–8 лет; опадая, она образует на поверхности
почвы толстый слой кислой подстилки, так как скорость ее минерализации довольно
низкая (8–12 лет). В наиболее благоприятных местообитаниях в лесной зоне – в
нижней части склонов гор, хорошо укрываемых зимой снегом, – горизонтально распростертые
по земле стволы кедрового стланика у основания могут достигать 30–40 см в
диаметре.
Синузия кедрового стланика на Северо-Востоке России
изменяется географически (с юга на север и от побережья вглубь континента) и
ландшафтно-экологически (с высотой над уровнем моря). В самых благоприятных
местах заросли кедрового стланика похожи на леса – это таежная темнохвойная
формация. В менее благоприятных местах по мере продвижения на север и ближе к
вершинам гор кусты становятся низкими и редкими, и здесь мы имеем дело со
своеобразной темнохвойной кустарниковой тундрой. Густота синузии
кедрового стланика зависит от глубины снегового укрытия.
Кедровый стланик в верхней части гор в лесном поясе, а также
на плакорах в зоне лесотундры и подзоне южной тундры может образовывать
сообщество совместно с синузиями зеленых мхов, кустистых лишайников и гипоарктических
кустарничков. Здесь он выступает как доминант и эдификатор. Кроме того, в
сочетании с синузией лиственницы, а нередко и каменной березы в Охотском округе
и березы плосколистной на Колыме, синузия кедрового стланика активно участвует
в сложении лесных сообществ. Отношения с лиственницей у кедрового стланика на
крутых склонах гор складываются так, что в ряде случаев он оказывается пионером
на вырубках и после пожаров. Спустя десятилетия, а иногда и столетия заросли
кедрового стланика в таких местах накапливают мелкозем, формируют или
восстанавливают эродировавшую после пожара почву и создают необходимые условия
для восстановления лиственничных лесов и редколесий.
Следует заметить, что экосистемы с развитой синузией
кедрового стланика имеют замедленный круговорот вещества; процессы накопления
органики (как в виде древесины, так и в виде хвои) преобладают над разложением.
В связи с этим кедровый стланик играет не последнюю роль в создании такой
ситуации, когда в ландшафте периодически накапливается огромное количество
быстро воспламеняющегося и хорошо горящего органического вещества. В этом
случае лесной пожар ускоряет возврат химических элементов в почву.
Симбиоз с корнями кедрового стланика образуют многие виды
грибов, в том числе и съедобные – белые и коричневые маслята, моховики, сыроежки.
Грибы наиболее обильны бывают во вторичных сообществах, где синузия кедрового
стланика сочетается с синузией березки Миддендорфа, зеленых мхов и кустистых
лишайников, а также отдельными особями возобновления лиственницы и каменной
березы. В таких местах в Охотском округе можно собирать и белые грибы. На
крутых склонах, обрывающихся в море, белый гриб не редок в Примагаданье и
хорошо известен грибникам.
Синузии деревьев.
Синузия
лиственницы (Larix cajanderi) вместе с
синузией кедрового стланика – основные ботанические элементы ландшафтов
Северо-Востока России. Ландшафтно-экологическая амплитуда этой синузии очень
велика от высокой поймы и надпойменных террас рек до олиготрофных сфагновых
болот и каменистых вершин сопок. Лиственница в нашем регионе – это основной
лесообразователь, и ее синузия геоботаникам и лесоведам хорошо известна как древостой,
или ярус деревьев в лесу. На север региона синузия лиственницы Каяндера
продвигается до р. Погынден, правого притока р. Мал. Анюй, на восток до р.
Майи, а вдоль Колымы лиственничники доходят почти до побережья
Восточно-Сибирского моря. В разных местообитаниях синузия лиственницы
сочетается со многими другими синузиями: кедрового стланика, березки
Миддендорфа, гипоарктических кустарничков, ксерофильных трав, мезофильных трав,
сфагновых мхов, зеленых мхов, кустистых лишайников и т.д. В связи с этим
геоботаниками и лесоведами выделяются разные типы лиственничных лесов:
зеленомошные, кустарничковые, лишайниковые, разнотравные, с подлеском из кедрового
стланика и т.д.
Для лиственничной
синузии характерно образование значительной ежегодно отмираемой фитомассы хвои,
которая, в отличие от хвои кедрового стланика, довольно быстро разлагается и
обеспечивает весьма интенсивный биологический круговорот вещества в суровых климатических
условиях. Таким образом, интенсивность биологического круговорота лиственничных
лесов значительно выше, чем зарослей кедрового стланика.
Общие запасы
древесины в лиственничных лесах в начале интенсивной колонизации края казались
высокими, однако это ложное мнение складывалось в основном из-за больших
площадей, занятых синузией лиственницы. В действительности же в большинстве
своем эти леса низкопродуктивны. После вырубки
деревья либо вообще не восстанавливаются, либо восстанавливаются за 800–1000
лет.
К сожалению, на большей части Магаданской области первичные
лиственничные леса были вырублены в 1930–1960-е годы во времена лагерной
колонизации Северо-Востока. Сегодня мы имеем здесь только молодые и вторичные
леса, мало-, а то и вовсе непригодные для эксплуатации на предмет заготовки
сырьевой древесины. Тем не менее, при расчете годичной лесосеки [годичная лесосека – это расчетное количество древесины,
которое подлежит ежегодной рубке на некоторой площади (район, область). Это
количество равно суммарному приросту древесины на этой площади в год.
Теоретически должно получаться, что вырубается древесины в области столько же,
сколько ее за это время прирастает] учитывались запасы и прирост древесины
не только в лесах, но и в редколесьях и рединах. Только этим в основном объясняется
тот факт, что расчетная лесосека в магаданской области в 1930-е годы составляла
около 5 млн. куб. м, а в последние 10 лет снижена до 1 млн 181 тыс. куб м. Фактическая же заготовка древесины во все годы оставалась по
сути на одном уровне – около 0,5 млн. куб.м в год. Даже при такой интенсивности
рубки площадь наиболее продуктивных долинных лесов в регионе ежегодно снижалась
на 50-100 кв. км. Всего с 1932 г. в Магаданской области заготовлено около 72
млн. куб м древесины.
С 1988 г. расчетная лесосека в области установлена в размере
362 тыс. куб. м. По всей вероятности, в связи с изменением экономики страны
развитие ситуации в лесном хозяйстве на Северо-Востоке России пойдет по другому,
нежели ранее, сценарию.
Синузия каменной березы (Betula ermanii) не имеет столь
широкого распространения, как синузия лиственницы Тем не менее это очень важным
компонент растительного покрова в приохотской части Магаданской области. Древостои
каменной березы встречаются здесь в ложбинах и на седловинах в горах – там, где
зимой скапливается много снега, но летом этот снег своевременно стаивает, и
нивальный режим выражен не очень сильно. Типичные местообитания каменной
березы в Приохотье вершины горных распадков, теплые склоны гор южной и
восточной экспозиций, а также склоны, обращенные к морю.
Для каменной березы характерно продуцирование большой
фитомассы листьев, относительно быстрый прирост древесины стволов и сучьев. О
быстром приросте стволов в толщину говорит, например, тонкий слой бересты у
этого вида березы, который снаружи постоянно лопается и обшелушивается. У
березы плосколистной береста отслаивается значительно медленнее, она прочнее и
толще.
Синузия каменной березы в растительном покрове может
сочетаться либо с синузиями кедрового стланика и зеленых мхов, либо с синузией
мезофильных трав, образуя в том и другом случае характерные для Приохотья
растительные сообщества. Надо сказать, что травяные каменноберезовые леса – это
экосистемы с наиболее интенсивным биологическим круговоротом. Здесь высока
продуктивность трав (местами до 100 ц/га сухой массы в год), листьев каменной
березы, ольхи кустарниковой, березки Миддендорфа. Скорость разложения опада
здесь самая высокая в ландшафте. Об этом можно судить, например, по тому, как
быстро перегнивают стволы березы по сравнению со стволами лиственницы и
кедрового стланика на гарях. Стволики березы среди валежника всегда имеют
степень разложения в 2–3 раза большую, чем стволы других дpeвесных.
Синузия чозении,
или ивы-кореянки (Choseniа arbutifolia) – один из самых
характерных и уникальных ценоэлементов в растительном покрове северо-Востока
России. Она образует своеобразные экосистемы в поймах рек и крупных ручьев всех
геоботанических округов, за исключением тундровой зоны. Внутриландшафтный же
ареал этой синузии очень узкий – за пределы поймы чозения не выходит. Места поселения
этого дерева – свежие галечники. Обильные побеги этой удивительной
породы появляются на галечниках одними из первых растений. Через год-два они
начинают сильно ветвиться у основания и образуют густую поросль, которая гасит
скорость одного потока и аккумулирует растительные остатки, песчаные и илистые
частицы, улучшая тем самым место своего обитания. Уже через 8–10 лет поросль
чозении похожа на молодой лес. Нижние сучья по мере роста отмирают, молодые
деревца быстро сбрасывают их и в своем росте резко устремляются вверх.
Тридцатилетняя синузия чозении – это древостой высотой до 20 м и с диаметром стволов
до 30 см. Галечник на этой стадии вокруг чозении уже сильно заилен,
сформирована примитивная почва, а под пологом деревьев неплохо развит
травянистый покров.
Зрелыми
насаждения чозении становятся в 50–60 лет, затем происходит их разрушение.
Древостой изреживается, многие особи усыхают. Если ранее в синузию чозении
внедрился тополь душистый, то по мере выпадения чозении он активизируется и
местами сменяет ее – правда, переживая ненадолго – всего на 40–50 лет.
Синузия чозении
обеспечивает интенсивный биогеохимический процесс за счет большого количества
образуемых каждый год листьев. К этому следует добавить очень обильное
семяношение, ежегодный отпад значительной биомассы в виде сучьев и коры. Ежегодный
прирост древесины у чозении столь интенсивен, что наружный слой коры постоянно
отшелушивается, так как быстро растущий в толщину ствол постоянно ее разрывает
и растрескивает.
Синузия
кустарниковых ив – это заросли высотой 1,5–2,5 (до 4,0) м. Кустарниковые виды
ив хорошо приспособленны к условиям пойменных и надпойменных террас – к
местообитаниям с весьма интенсивным биологическим круговоротом. При относительно
небольшой фитомассе стволов и сучьев синузия кустарниковых ив ежегодно
производит значительную фитомассу листьев – до 3–5 т/га сырой массы в зоне гипоарктических тундр и до 8–10 т/га
сырой массы в лесной зоне.
Ивы ежегодно и
обильно плодоносят, хорошо поедаются лосем и северным оленем, образуют дополнительные экологические ниши для многих видов
животных. Совместно с синузиями мезофильных трав, гипновых и зеленых мхов синузия
кустарниковых ив образует многоярусные сообщества с очень высокой
продуктивностью. В биогеохимическом плане эта синузия сходна с синузией
чозении.
Разные виды ив несколько отличаются своей экологией и
географией. На галечниках и пляжах растут Salix alaxensis, Salix sсhwerinii; там, где аллювиальный процесс ослаблен, синузию
образуют Salix lanata, Salix kolymensis, Salix udensis, а вне долин рек по ручьям и слегка заболоченным ложбинам стока
на склонах и шлейфах склонов встречаются Salix krylovii, Salix bebbiana, Salix
pulchra и др. При этом Salix alaxensis распространена только на Чукотке.