Вы находитесь: НАША БОТАНИЧКА — Теория — Биоморфология — Антропова Г. Л. Биоморфология розоцветных Северо-Востока СССР
ОНТОГЕНЕЗЫ БИОМОРФ ИССЛЕДОВАННЫХ ВИДОВ (ПРОДОЛЖЕНИЕ)
РЯБИННИК ПАЛЛАСА SORBARIA PALLASII (D. DON)
POJARK.
На Северо-Востоке СССР этот вид встречается достаточно
редко, отмечен в Колымском флористическом районе [Хохряков, 1985]. В прилежащих
районах обычен на Камчатке, Сахалине, в Приморье [Ворошилов, 1982], в южных
частях Якутии.
Исследования проводились на границе Магаданской области и
Хабаровского края в верховьях р. Кулу в 1938 г. Рябинник растет на мелко- и
крупнощебнистых склонах сопки на высоте 35 м над ур. м. Популяция занимает
небольшой участок около 8 м2 на мохово-лишайниковом склоне. Рядом растут виды,
характерные для подобных мест обитания: Ledum palustre L., Vассinium vitis-idaea L., Vассinium uliginosum L., Cassiope
ericoides (Pall.) D. Dоn., Spiraea stеvеnii L. (Sсhnеid.).
Проростки и молодые экземпляры в популяции не обнаружены. Это связано с тем,
что семенное возобновление в популяции затруднено, так как участок достаточно
густо заселен другой растительностью (если и встречаются свободные пятна, то и
они покрыты моховыми и лишайниковыми латками). К тому же данное местообитание
вида расположено на северном пределе его ареала. Здесь растение хорошо
возобновляется вегетативным путем, молодые растения быстро теряют стержневой
корень, и, формируя придаточную корневую систему, переходят к вегетативно-подвижному
образу жизни, размножаясь с помощью гипогеогенных корневищ, используя любой
более или менее свободный участок склона для образования парциальных кустов.
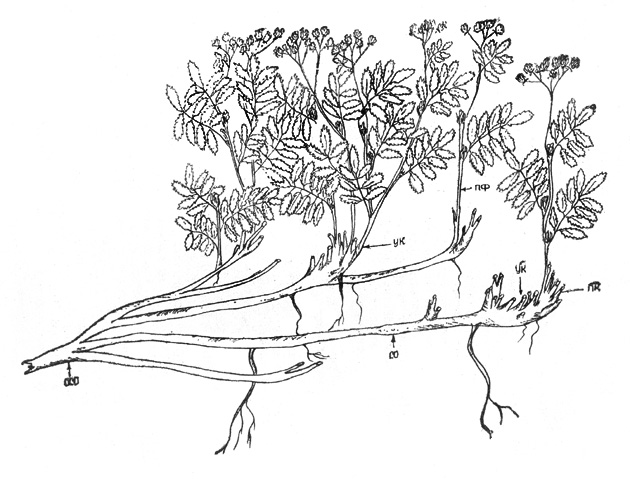
Рис. 33. Рябинник Палласа. Часть основной скелетной оси
|
Подземная система органов представлена гипогеогенными
горизонтально нарастающими корневищами и системой придаточных корней, которые.
обычны и на корневище, и в узле кущения парциальных кустов. Корневище отрастает
от подземной части куста. Длительное время растет плагиотропно на глубине 5-8
см. По всей его длине сидят многочисленные почки, которые некоторое время
находятся в состоянии покоя. Особенно много их на молодых частях корневища. На
старых участках большинство почек отмирает, а те немногие, что остаются живыми,
постепенно увеличиваются в размерах до 3-4 мм, покрываются плотными чешуями и в
таком виде способны к длительному существованию в состоянии покоя. Спящие почки
со временем реализуются либо в новые подземные, либо в надземные ортотропные
побеги формирования, которые становятся первым звеном в развитии парциального
куста.
По всей длине корневища располагаются придаточные корни,
однако они в основном ослаблены и редки. Основная масса этих корней
сосредоточена на молодых частях корневища; они образуются ежегодно, но ежегодно
и отмирают, лишь некоторые из них в более старых частях корневища остаются
живыми и нарастают в течение нескольких лет.
На базе корневищ формируется система подземной скелетной
основы растения. Со временем подземные скелетные оси (корневища) утолщаются.
Так, если молодые одногодичные корневища имеют толщину около 1 мм, то старые
многолетние - до 1 см. Основная скелетная ось (старое многолетнее корневище)
образует разветвленную систему подземных СО, растущих в разных направлениях.
Длина отдельных СО достигает 3-4 м и формирует 8-10 порядков боковых дочерних
осей.
Практически данная популяция представлена одной разросшейся
куртиной, которая еще не приобрела статус клона, так как все парциальные кусты
и отдельные надземные ПФ связаны между собой подземными осями. Образование
парциальных кустов начинается выходом на поверхность верхушки подземного
побега, до этого нарастающего горизонтально. Бывает, что первые надземные
побеги формируются из спящих почек, расположенных более или менее равномерно по
всей длине СО, однако чаще они реализуются в побеги, развивающиеся по типу ПД
и, как правило, большинство их оказываются эфемерными. ПФ 1-го года 4-5 см
длиной с 2-3 очередными листьями. Листья непарноперисто-сложные, с 5-6 парам и
долей, каждая из которых 3-4 см длиной. Междоузлия 1,5-2 см. В пазухе каждого
листа сформированы хорошо видимые почки. В дальнейшем с увеличением размера и
порядка ПФ почки увеличиваются до 3-4 мм. К концу вегетационного периода ПФ
отмирает до второго или даже до первого узла от его основания, т. е.
жизнеспособными сохраняются 1-2 перезимовавшие почки; из них и развиваются ПФ
следующего года. |
Размер последнего обычно несколько больше размера побега
предыдущего года, и он также отмирает к концу вегетационного периода. От
отмерших побегов остаются пеньки с живыми почками, которые дают начало новым ПФ
более высокого порядка. Иногда отмирание ПФ бывает более полным: до уровня
подстилающей поверхности или до корневища. Тогда новый побег формирования
образуется из спящей почки корневища, наиболее близко расположенной к отмершему
побегу. Такой побег может сразу приобретать ортотропность или некоторое время
растет горизонтально в почве и лишь потом меняет направление роста на
восходяще-ортотропное, выходит на поверхность и в дальнейшем проходит цикл
развития, аналогичный предыдущим ПФ.
Постепенно длина ПФ увеличивается сначала до 6-8 см, потом,
к 6 годам, достигает максимального размера - 10-12 см. Одновременно
увеличивается размер (до 5-6 см) и количество (до 3-4) листьев на нем. С этого
времени у р. Палласа начинается формирование репродуктивных побегов. Цветонос
терминального типа, развивается как продолжение вегетативного побега, причем в
основании его находится лист с крупной (7 х 7 мм) вегетативной почкой, которая
занимает положение между листом и репродуктивным приростом и визуально
воспринимается как терминальная, хотя в действительности является обычной
пазушной почкой.
Репродуктивный прирост отличается от вегетативного.
Во-первых, он меньшего диаметра от самого основания и потому хорошо от него
отличим. Во-вторых, он несет меньшее число листьев, они меньших размеров и
верхние редуцированы более, чем нижние; на вегетативном же приросте все листья
приблизительно одинакового размера. В-третьих, пазушные почки репродуктивного
побега значительно. меньше - всего около 1 мм. Сам цветонос по длине лишь
незначительно уступает материнскому и составляет 8-12 см. Заканчивается
цветоносный побег головчатым соцветием, состоящим из 6-8 мелких цветков. Таким
образом, теперь весь надземный ортотропный побег состоит из двух приростов
-вегетативного и репродуктивного, возникших за один сезон и составляющих вместе
20-25 см. Это предельная высота растения в данном местообитании.
Нижняя часть оснований побегов формирует узел кущения.
Пенечки, оставшиеся от отмерших побегов, порой так близко прилегают друг к
другу, что могут срастаться, некоторые остаются отдельно, и тогда узел кущения
по структуре напоминает каудекс. Располагается узел кущения на уровне
подстилающей поверхности так, что нижняя его часть скрыта под ней, а верхняя
незначительно (на 1-3 см) возвышается сверху. От верхней части отрастают
вегетирующие и плодоносящие побеги, от нижней - новые корневища. Надземные
побеги чаще всего не ветвятся. Лишь иногда из основания отмершего ПФ сразу две
зимующие почки трогаются в рост. Но нарастают они неодинаково. Одна из них дает
обычный удлиненный ПФ, который развивается по той же схеме, что и предыдущие
побеги, другая дает укороченный побег длиной около 1 см. Последний к концу
сезона формирует терминальную почку, из которой на следующий год вырастает
новый укороченный побег, а из пазушной почки его отрастает удлиненный ПВ,
аналогичный ПФ. И только на 3-й год терминальная почка дает удлиненный прирост,
заканчивающийся соцветием. Однако такой 3-летний цикл развития не совсем
типичен для р. Палласа. Абсолютное большинство его ПФ заканчивают цикл развития
в течение одного сезона (рис. 33).
Цикл развития отдельного парциального куста продолжается не
более 12-15 лет. Репродуктивная активность длится 6-8 лет, затем она падает,
после чего начинается деградация куста, которая через 3-4 года заканчивается
его отмиранием. Большая часть парциальных кустов исследуемой популяции
представлена молодыми и средневозрастными кустами, но иногда встречаются
стареющие и отмирающие. у стареющего куста развит достаточно массивный каудекс,
однако в большей своей части уже не функционирующий. Редкие побеги,
появляющиеся на нем в текущем году, тонкие, ослабленные, не более 5 см длиной,
отмирающие к концу сезона. Через 2-3 года новые побеги перестают появляться -
куст отмирает.
По данным М. Т. Мазуренко и А. П. Хохрякова [1976], в более
благоприятной обстановке (пос. Хениканджа Тенькинского района) в долине ручья
р. Палласа образует высокие кусты от 40 см до 1 м. И хотя его ПФ также отмирают
на значительную длину, многолетние основания побегов могут достигать 20-30 см,
они обладают способностью полегать и укореняться. От укоренившейся части
обильно отрастают ПФ. Основной цикл развития равен 10-12 годам. Становится
ясно, что р. Палласа имеет по крайней мере две экобиоморфы: одна - кустарник, с
циклом развития в 10-12 лет; другая - кустарничек, с признаками, приближающими
его к травянистым многолетникам, о чем свидетельствует минимальный цикл
развития его побегов, равный 1 году.
Таким образом, в пределах этого вида прослеживаются
изменения в структуре биоморф, направленных в сторону преобразования
кустарниковых форм роста в кустарничковые с элементами полукустарников, о чем
свидетельствует активное и глубокое отмирание побегов. При ухудшении
климатической и экологической обстановки это может привести к образованию длиннокорневищных
травянистых многолетников.
ДРИАДА БОЛЬШАЯ DRYAS GRANDIS JUZ.
Дриады - стелющиеся растения, с полностью распростертыми
скелетными осями и побегами. Для обозначения их жизненной формы используются
разные названия: по определению Варминга [Warming, 1909], шпалерные
кустарнички, в классификации И. Г. Серебрякова [1962] - вегетативно-подвижные
кустарнички шпалерного типа. Мы используем термин «стланички» [Юрцев, 1968].
Изучено 4 вида дриад. Дриада большая - голарктический вид, встречается по
галечникам и известняковым отложениям Восточной Сибири: Ленско-Колымский,
Ангаро-Саянский флористические районы; отдельные находки зарегистрированы в
Северной Монголии [Юзепчук, 1941]. В пределах изучаемого региона обычен в континентальных
районах: Охотско-Колымском водораздельном, Охотско-Омолонском водораздельном,
Омолонско-Анюйском. Реже он встречается в Чаунско-Центрально-Чукотском и
Средне-Чукотском флористических районах.
Материал собран в континентальных районах Магаданской
области в 1972-1979 гг. в следующих пунктах: в Северо-Эвенском районе - в
окрестностях перевалочной базы «Кегали» и в пойме р. Хадаранджа; в Билибинском
районе - в окрестностях перевалочной базы «Уляшка»; В Тенькинском районе - в
окрестностях пос. Кулу и на стационаре «Контакт»; В Сусуманском районе - в
устье р. Урультун; на границе Магаданской области и Хабаровского края - в
верховьях р. Кулу.
Выделено три типа условий произрастания вида: первый тип - обычные
или средние - разреженные долинные ерники на галечнике (пойма р. Авланди);
второй - оптимальные - лиственничные редколесья по долинам рек и ручьев (устье
р. Урультун); третий - экстремальные - приподнятые террасы горных рек и ручьев
(верховье р. Кулу). Этим типам местообитаний соответствуют три экологические
формы (экобиоморфы) д. большой. Ниже приводится описание экобиоморфы 1 по
материалам, собранным в пойме р. Авланди (окрестности перевалочной базы
«Кегали») среди разреженного долинного ерника- средний тип условий произрастания.
Остальные две экобиоморфы сравниваются по линейным пара метрам органов и
изменениям в структуре побегов.
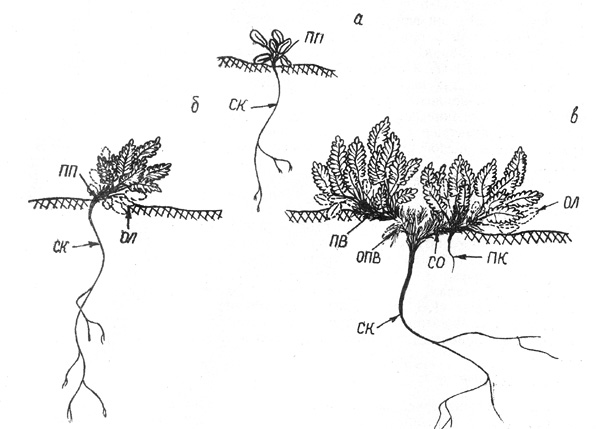
Рис. 34. Дриада большая. Возрастной ряд особей: а -
проросток, б - 3-й год (полегание ПП), в - б-й год (ветвление ПП). Здесь и на
рис. 35, 39 ОЛ - отмершие листья, ОПВ - отмершие ПВ
|
Экобиоморфа 1
На галечнике среди зарослей и отдельных кустов березы тощей
рассредоточены куртины д. большой. Растения находятся в условиях обедненных
почв, но достаточного увлажнения. Защищены от прямого ветрового воздействия.
В онтогенезе экобиоморфы 1 выделено 4 фазы: 1) проросток
(1-й год жизни), 2) нарастание ПП (2-3-й годы), 3) развитие системы ПП (4-7-й
годы), 4) формирование скелетных осей, активный рост их побегов (8-35-й годы),
5) жизнь в виде клона (с 35 лет).
Пер в а я ф а з а. Семена прорастают без периода покоя. В
конце июля-начале августа рядом со взрослыми плодоносящими растениями
появляется большое число проростков. Проростки выносят на поверхность семядоли,
вслед за которыми развиваются 3-5 настоящих листьев размером до 3 мм. К концу
1-го года вегетации ПП имеет длину 1-1,5 см, ортотропный. Длина корня 3 мм
(рис. 34, а).
В т о р а я ф а з а. В течение последующих 2-3 лет
продолжается нарастание ПП, его годичные приросты по 3-5 мм. Надземная часть ПП
увеличивается незначительно, так как его основание присыпается листовым опадом,
частичками почвы. На 3-й год основание ПП несколько изгибается, но полностью
еще не полегает. Корень увеличивается до 7-9 см. На годичных приростах
развивается по 5-7 листьев, сближенных на верхушке, размером по 3-7 мм.
Прошлогодние листья желтеют, но не опадают, а остаются на стебле, предохраняя
побег и пазушные почки от вымерзания (рис. 34, б).
Т р е т ь я ф а з а. На 4-5-й год ПП моноподиально
ветвится. 1-3 укороченных ПВ формируются из зимующих почек прироста предыдущего
года. На следующий год, также моноподиально, появляются ПВ второго порядка, в
числе 6-8. Некоторые из них ослабленные и после 3-6 лет роста отмирают,
остальные (5-7) продолжают нарастание. ПВ имеют восходяще-ортотропное
направление роста. Образуется система ПП. С увеличением возраста растения ПП
перестает выделяться среди ПВ: и длина, и толщина их приблизительно равны.
Растущие ПП и ПВ составляют СО, которые участвуют в дальнейшем формировании
жизненной формы растения. Количество СО может быть разным - от 3 до 5.
Основания побегов полегают, вертикальными остаются приросты 1-2 последних лет
(рис. 34, в). |
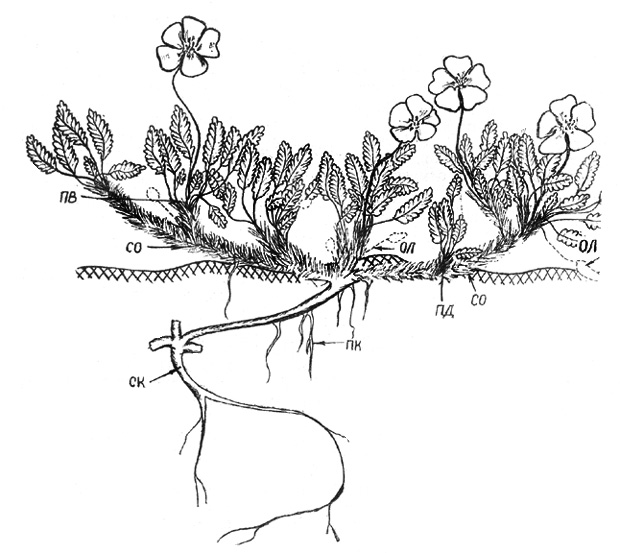
Рис. 35. Дриада большая. Одна из основных скелетных осей
|
Ч е т в е р т а я ф а з а. Начиная с 8-го года наиболее
развитые СО дают репродуктивные побеги, они неразветвленные с единичным
терминальным цветком. Цветоножка в момент распускания цветка имеет размер 3-4
см, к концу плодоношения вытягивается и достигает 18 см. Б. А. Тихомиров [1963]отмечал,
что подобное явление - цветение растений при минимальных размерах цветоносного
побега - свойственно многим арктическим видам. Одновременно с репродуктивным
побегом из пазушных почек листьев предыдущего года образуется 3-6 замещающих ПВ
(рис. 35). Часть из них, не зацветая, отмирает уже к следующему году, некоторые
- через 1-2 года, и лишь 1-2 побега продолжают нарастание СО. В последующие
годы происходит увеличение числа репродуктивных побегов. Замещающие ПВ,
образовавшиеся под ними, после 2-3 лет роста, в свою очередь, развивают цветок
и розетку замещающих ПВ следующего порядка. К 15-летнему возрасту все побеги
становятся ди-трициклическими. Происходит замена моноподиального типа ветвления
симподиальным. Однако замещающие ПВ в промежутке от своего появления до
зацветания могут развивать ослабленные недолговечные боковые побеги, которые
отмирают в течение 1-3 лет. Иногда они могут формировать репродуктивные побеги,
но после цветения отмирают полностью. Таким образом, наряду с симподиальным
ветвлением сохраняется моноподиальное, т. е. у д. большой ветвление моноподиально-симподиального
типа (рис. 36). |
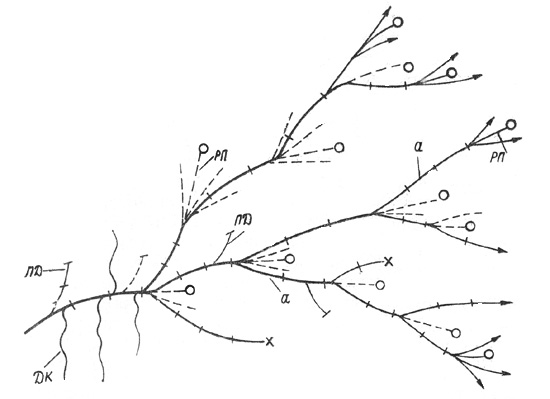
Рис. 36. Дриада большая. Схема ветвления побегов: а -
замещающий ПВ
|
На 10-15-й год в разных частях побегов из спящих почек
начинают появляться ПД. Большинство из них эфемерные, но некоторые растут до
7-8 лет, ветвятся, формируя СПД. После цветения, если оно происходит, система
побегов дополнения отмирает. ПД, выполняя роль побегов обогащения, увеличивают
ассимиляционную поверхность растения, кроме того, после отмирания они
продолжительное время (как и листья) сохраняются на материнском побеге, защищая
его от неблагоприятных воздействий. Постепенно происходит увеличение приростов
побегов до 1,5-2 см, поэтому к 15-летнему возрасту СО, образованные несколькими
замещающими побегами ветвления, хорошо развиты: длиной 15-18 см, толщиной у
основания - 3,5 мм. Растение выглядит округлой зеленой латкой 30-36 см в
диаметре, распластанной по поверхности субстрата. Приподнимающиеся части
побегов около 4 см в высоту. Это приросты 2 последних лет. Две трети каждой СО
лежит на поверхности, остальная (базальная) часть погружена в почву. В
основании СО образуются придаточные корни. С момента их возникновения СО
перестают утолщаться, способность ко вторичному росту сохраняет лишь привершинная
часть побегов. Подобная закономерность была отмечена ранее исследователями
стланиковых жизненных форм [Полозова, 1978; Колищук, 1967; Хохряков, 1976б, и
др.].
П я т а я ф а з а. За прошедший период шло накопление
вегетативной массы растения, преобладали процессы нарастания, формировались СО,
системы побегов: СПП, СПВ, СПД. Отмирали лишь ослабленные ПВ и эфемерные ПД.
С 15-20-летнего возраста процессы нарастания-отмирания
уравновешиваются, растение достигает стабильного состояния. Число ежегодно
появляющихся побегов приблизительно равно числу отмирающих. Подсчитано, что на
одной из модельных ветвей (примерно 18-летнего возраста) вновь появилось 12 ПД
и 32 ПВ, отмерло в этом же году 10 ПД и 28 ПВ, на другой (того же возраста) -
соответственно 19 ПД и 36 ПВ, отмерло - 15 ПД и 33 ПВ. В среднем из подсчета на
10 модельных ветвях в. период четвертой фазы ежегодно появляется 48 побегов и
отмирает 46. |
К 20-летнему возрасту куртина стланичка имеет диаметр 50-60
см. Мощный стержневой корень толщиной у основания 1-1,5 см; от него отходят 5-7
СО длиной 25-30 см. Придаточные корни образуются только на тех частях СО,
которые погружены в почву. Та их часть, которая лежит на поверхности,
придаточных корней не образует совсем или образует очень редко. В этот период
растение имеет до трех-четырех порядков ПВ.
У растений старше 35 лет усиливается отмирание побегов. Базальные
части СО уже не имеют вегетирующих побегов, все ПД, образовавшиеся ранее,
отмирают, новые здесь не образуются. Ослабленные ПВ также отмирают, остаются
немногочисленные СО, покрытые отмершими побегами и листьями. Ветвящиеся части
СО все более удаляются от основания корня, их базальные части постепенно
оголяются, разрушаются, и они теряют связь с корнем. Куртина распадается -
отдельные СО обособляются и продолжают существовать самостоятельно, нарастая
верхушкой и одновременно отмирая с базального конца. Растение с этого времени
существует в виде клона. Питание и прикрепление к субстрату ему обеспечивает
система придаточных корней, Характерным для д. большой является то, что СО ее в
данном местообитании не расходятся на большое расстояние друг от друга и всегда
можно определить границы клона. Предельная величина его, отмеченная нами, -
130-150 см в диаметре.
Постепенно все более проявляются признаки старения у
растений. Уменьшаются годичные приросты побегов до 3-5 мм. СО продолжают
отмирать с базального конца с нарастающей интенсивностью. Более разреженной
становится зона ветвления побегов. Большинство появляющихся ПВ и ПД отмирает в
этом же году. В дальнейшем происходит отмирание и отдельных систем побегов.
Продолжительность жизни д. большой в данном местообитании в среднем составляет
60-75 лет.
Экобиоморфа 2
Устье р. Урультун. Оптимальные условия произрастания вида:
почва богата гумусом, достаточно хорошо увлажнена, зимой глубокий снежный
покров надежно защищает растения от вымерзания, летом растения защищены от
воздействия ветра растущей рядом растительностью (лиственницей, кустарниковыми
видами ив).
Размеры растений по сравнению с экобиоморфой 1 значительно
увеличены: максимальная величина годичных приростов достигает 3,5-4 см,
листовой пластинки - 5,5 см, цветоноса - 22 см. Приподнимающиеся части побегов
в высоту 6-8 см, это приросты 2-3 последних лет. К концу онтогенеза СО, как
правило, не теряют связь со стержневым корнем, который существует до конца
онтогенеза. Сходные данные получены М. Т. Мазуренко и А. П. Хохряковым [1977]
для растений из района оз. Дарпир (Момский район Якутской АССР). Они указывают,
что первичная ось (у нас ПП) д. большой нарастает в течение 5-10 лет и
достигает 8-12 см в высоту, ортотропные части восходящих СО равны 5 см и
состоят из 3-5 розеточных приростов. Годичные побеги в зависимости от условий
имеют длину от 2 до 5 см.
Экобиоморфа 3
Встречается в пойме р. Авланди и в верховьях р. Кулу.
Обдуваемые ветром приподнятые приречные террасы с каменистым субстратом и со
слабо выраженной растительностью или галечные отмели в пойме -реки,
периодически затопляемые паводковыми водами. Условия экстремальные для биоморф
д. большой.
Годичные приросты побегов имеют длину 5-6 мм, листья - 1-1,2 см, цветоносы - 6 см. Над субстратом растения возвышаются на 1,5-2 см.
Благодаря интенсивно образующейся придаточной корневой системе СО стланичка
отходят на большое расстояние от стержневого корня и значительно раньше, чем в
предыдущих случаях, теряют с ним связь. Характерно то, что ветви одного клона
перекрываются ветвями другого, поэтому выявить границы отдельного клона
практически невозможно. Образуется сплошной ковер из многих особей. Иногда на
ранних этапах онтогенеза вследствие каких-либо причин может произойти отмирание
ПП. В этом случае в базальной части ПП из пазушной почки, которая находилась в
состоянии покоя 2-3 года, развивается побег формирования. В дальнейшем он
играет основную роль в формировании стланичка, так как замещает ПП. Приросты ПФ
в первые 1-2 года достигают 2,5 см, междоузлия несколько растянуты, листья на
нем распределены равномерно. На 3-й год он ветвится, и в дальнейшем развитие
СПФ не отличается от развития СПП.
Таким образом, в нарастании систем побегов происходят
изменения в ряду экобиоморф от оптимальных условий произрастания к
экстремальным. На первых этапах онтогенеза нарастание побегов идет однотипно,
различия затрагивают только линейные размеры органов, в дальнейшем происходят
более глубокие изменения. У экобиоморфы 2 (оптимальный тип местообитаний)
стержневой корень сохраняется в течение всей жизни, придаточное укоренение
выражено достаточно слабо, биоморфа представляет собой кустарничек с
приподнимающимися до 6-8 см в высоту побегами. У экобиоморфы 1 размеры органов
меньше, над субстратом растение приподнимается на 4 см. К концу онтогенеза она
приобретает частичную вегетативную подвижность, так как теряется связь СО друг
с другом. У экобиоморфы 3 высота приподнимающихся частей побегов не более 2 см,
имеет место более активное укоренение побегов и вследствие этого более
длительное существование огделившихся скелетных осей растения. Из сказанного
следует, что в ряду этих экобиоморф происходит усиление вегетативной
подвижности растений и стремление к большей геофитизации побегов.
ДРИАДА ТОЧЕЧНАЯ DRYAS PUNCTATA JUZ.
Дриада точечная - почти циркумполярный (с дизъюнкцией в
Центральной и Восточной Канаде, Западной и. Северной Гренландии)
арктоальпийский вид [Юрцев, 1984]. На территории Северо-Востока СССР
характеризуется массовым распространением везде, кроме Прибрежно-Охотского
флористического района, редка в Гижигинском районе.
Материал собирался в течение ряда лет в разных пунктах
Магаданской области. Выделено два типа условий местообитания вида: 1) горные
тундры континентальных районов области, здесь д. точечная занимает большие
пространства, иногда в десятки гектаров; 2) приморские склоны, подверженные
действию сильных ветров в течение всего года (зимой склоны обнажены, снег почти
полностью выдувается).
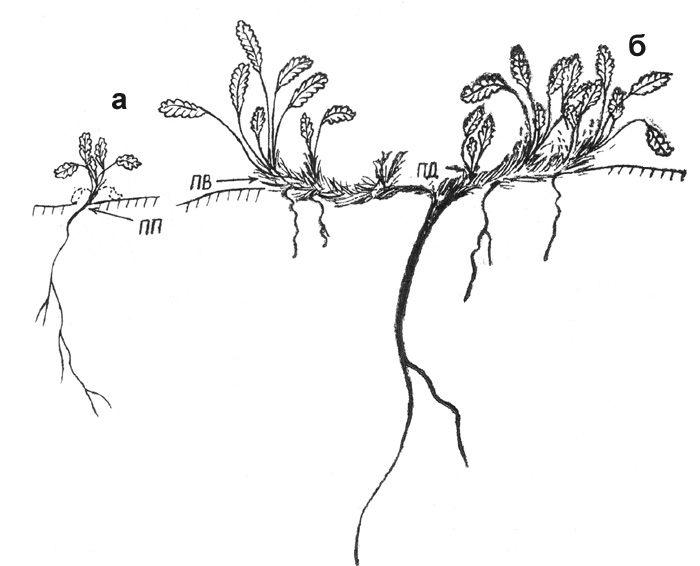
Рис. 37. Дриада точечная: а - проросток, б - первичный куст
|
Экобиоморфа 1
Ниже приводится описание онтогенеза д. точечной, растущей в
окрестностях стационара «Контакт». Исследования проводились в течение полевых
сезонов 1977, 1983, 1986 гг.
В онтогенезе д. точечной выделено 4 фазы: 1) проросток (1-й
год), 2) формирование СПП (2-7-й годы), 3) вычленение основных скелетных осей
(7-20-й годы), 4) жизнь в виде клона (с 20 лет).
П е р в а я ф а з а. Проросток д. точечной аналогичен
проростку д. большой - это укороченный побег розеточного типа (рис. 37).
В т о р а я ф а з а. На 2-й год терминальная почка ПП
образует новый прирост, прошлогодний же к концу текущего вегетационного периода
полегает. ПП начинает моноподиально ветвиться, давая 1-3 (5) ПВ. На приростах
раскрывается от 3 до 10 ассимилирующих листьев. На 4-й год также моноподиально
образуется еще 5-7 ПВ. Одни из них угнетены и вскоре отмирают, другие
продолжают нарастание, участвуя в формировании скелетных осей стланичка. Через
2-3 года основания ПВ полегают, ортотропными остаются приросты 1-2 последних
лет от 0,7 до 1,2 см высотой. На 5-6-й год в основании побегов из спящих почек
развиваются ПД.
Т р е т ь я ф а з а. У 7-15-летних растений происходит
вычленение СО. Обычно ПВ образуется много (7-8), но так как нормальный рост и
развитие приобретают 2-3 из них, то остаются в живых немногие побеги, которые
являются составной частью формирующихся СО. Последние представляют собой скелет
растения. На 7-8-й год в основании полегших побегов, присыпанных почвой,
развиваются придаточные корни, через которые растение получает дополнительное
питание и которые, кроме того, способствуют погружению побегов в почву.
Годичные побеги увеличиваются в размерах с 4-5 до 7-8 мм. В
11-12 лет д. точечная дает первые репродуктивные побеги, под которыми
образуется 7-8 замещающих ПВ. Те из них, которые в 1-й год усиливаются,
замещают материнский побег, нарастая по направлению его роста, остальные
отмирают. Происходит смена моноподиального типа ветвления симподиальным. |
К 16-летнему возрасту стланичек обильно цветет, обильно
плодоносит, под каждым репродуктивным побегом образуется большое количество
замещающих (рис. 38). Наступает период стабилизации - процессы
нарастания-отмирания уравновешиваются. Зона ветвления побегов удаляется от
стрежневого корня. Обеспечение минеральным питанием приходится на систему
придаточных корней, которые следуют за молодыми частями побегов и обычны на
приростах 7-8 последних лет и старше. Растет число ПД, они появляются в разных
частях СО. Благодаря им увеличивается ассимиляционная поверхность растения и
его защищенность от неблагоприятных воздействий среды. К 20-летнему возрасту
стланичек выглядит округлым зеленым пятном 50-60 см в диаметре.
Ч е т в е р т а я ф а з а. Постепенно стержневой корень
утрачивает ведущую роль в обеспечении растения питанием и вскоре разрушается,
следом разрушаются и базальные участки СО, в результате чего происходит их
обособление. Таким образом, растение продолжает существовать уже в виде клона.
Установить предельный возраст д. точечной невозможно, так как клон может
существовать неопределенно долгое время. Отдельные системы побегов, продолжая
нарастать верхушкой, отмирают с базального конца. Как и у экобиоморфы 3 д.
большой, ветви одного клона перекрываются ветвями другого. В дриадовых тундрах
бывает невозможно установить принадлежность обособившихся частей растений к
определенному клону. Однако отдельные СО имеют возраст до 40 лет и более
(возраст установлен по подсчету
среднегодичных приростов побегов). Длина их в этом возрасте 60- 70 см. Все
годичные побеги старше 7 лет укоренившиеся. Придаточные корни в момент
возникновения волосовидные; по мере роста удлиняются, разветвляются, у
основания могут иметь до 1 мм в диаметре.
Более мощных корней д. точечная не образует (рис. 38).
Экобиоморфа 2
Растения этой экобиоморфы наблюдались в 1976 г. в
окрестностях пос. Хатырка Беринговского района Магаданской области. Д. точечная
растет здесь на оголенных слабозадернованных приморских склонах. В течение
всего года подвержена постоянно дующим здесь сильным ветрам. В зимний период
снег почти полностью сдувается со склонов, летом ощущается дефицит влаги. Почва
слабо сформирована, щебнистая. Условия крайне экстремальные для жизни растений.
В отличие от описанных выше растений, у
экобиоморфы-2-размеры годичных побегов, листьев, черешков, репродуктивных
побегов значительно меньше. Ортотропные части побегов возвышаются над субстратом
всего на 2-3 см, но наиболее характерно, что к концу вегетационного периода
побеги полегают полностью, даже их верхушки плотно прижимаются к субстрату. Все
отмершие листья и большинство побегов сохраняются на стеблях, плотно окутывая-
их и предохраняя от вымерзания в суровый зимний период. Придаточные корни
типичны для приростов 3-4 последних лет - они активно следуют за молодыми
частями побегов. |
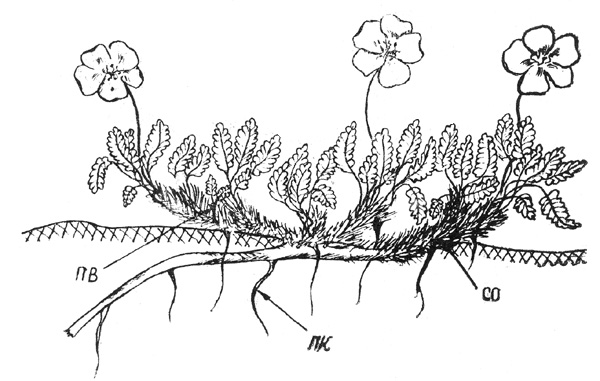
Рис. 38. Дриада точечная. Часть основной скелетной оси |
ДРИАДА ЦЕЛЬНОЛИСТНАЯ DRYAS INTEGRIFOLIA VAHL.
Дриада цельнолистная - чукотско-североамериканский вид. Подробный
ареал представлен в «Арктической флоре. СССР» [1984]. На исследуемой территории
встречается в северных частях региона, включая о-в Врангеля, достаточно редка в
Колымском флористическом районе.
Описание онтогенеза велось по материалам, собранным в Сусуманском
районе Магаданской области, в устье р. Урультун в 1978 г. Здесь д.
цельнолистная тяготеет к приручьевым террасам, реже поселяется на щебнистых
осыпных склонах. В онтогенезе выделено 4 фазы: 1) проросток (1-й год жизни),
2) формирование СПП (2-15-й годы), 3) вычленение основных СО (15-25-й годы), 4)
жизнь в виде клона (с 25 лет).
П е р в а я ф а з а. Проросток ортотропный, розеточный,
несет 7-9 листьев. Стержневой корень 3-5 см, слабо разветвлен на верхушке. К
концу первого вегетационного периода основание ПП полегает, лишь верхушка
побега сохраняет ортотропность.
В т о р а я ф а з а. На следующий год нарастание ПП
продолжается, его прирост 2-4 мм, на нем 10-13 листьев, сближенных на верхушке
побега. Ветвление ПП начинается на 2-3-й год, образуется до 6 ПВ первого
порядка. Приросты ПВ такие же, как у ПП, - 2-4 мм. Большинство их отмирает в 1-й
же год, лишь 1-3 продолжают рост. Следующие 1-2 года образовавшиеся побеги
нарастают не ветвясь. В это время из оснований нарастающих побегов трогаются в
рост спящие почки, которые дают ПД. Иногда эти побеги приобретают усиленный рост,
длина их прироста может достигать 1,5-2 см. Однако в сравнении с ПВ они тонкие
и после 1-3 лет роста отмирают. ПП на 4-5-й год ветвится вторично, в это же
время ветвятся и растущие ПВ. Частички почвы, опавшие листья, побеги, слоев ища
лишайников скапливаются в основании побегов, благодаря чему последние
оказываются скрытыми в подстилке. Здесь на 6-7-й год образуются придаточные
корни.
В течение последующих лет развитие побегов продолжается так
же, как и раньше, т. е. ослабленные побеги отмирают, более сильные ветвятся
через каждые 1-2 года, участвуя в дальнейшем становлении жизненной формы
растения. Величина годичного прироста возрастает до 1 см, но по-прежнему
ортотропной остается лишь верхушка побега, приподнимающаяся на 3-4 см над
почвой.
Т р е т ь я ф а з а. К 15-летнему возрасту стланичек
разрастается, у него вычленяется 5-7 основных СО, которые радиально расходятся
от основания стержневого корня. К этому времени они достигают размера 10- 12
см. Однако толщина их почти на всем протяжении одинакова (2 мм), лишь у
основания несколько больше (3 мм). Это свидетельство того, что вторичное
утолщение побегов осуществляется непродолжительное время - до момента появления
придаточных корней. Репродуктивные побеги развиваются на 15-17-й год, под ними
образуется большое число (8-10) замещающих ПВ, но лишь 2-3 из них становятся
многолетними, сохраняют нормальный рост и развитие. К этому времени характерно
более активное появление ПД. Они, как правило, ортотропны и живут не более 1
года.
Особенностью д. цельнолистной является то, что некоторые из
ПВ, растущих ближе к верхушке материнской оси, приобретают усиленный рост, и их
приросты могут достигать 3 см. Чаще всего это случается у растений, обитающих
на хорошо дренированных почвах приручьевых склонов. Но не всегда именно эти
побеги становятся продолжением материнской оси, они часто отмирают, так как
ослаблены и этиолированы - 0,3-0,5 мм в диаметре. Но бывает, что они
укореняются в основании и тогда намного опережают в росте ПВ и ПД, появившиеся
с ними одновременно. В этом случае именно они продолжают нарастание скелетной
оси. Латка стланичка теряет округлую форму, вытягиваясь в сторону нарастания
этих побегов. По всей видимости, эти побеги с усиленным ростом являются
аналогами побегов формирования.
Ч е т в е р т а я ф а з а. В 20-25 лет растение обильно
цветет и плодоносит, активно образует многочисленные придаточные корни уже на
приростах 4-5 последних лет, и не только в тех частях, которые скрыты в почве,
но и в тех, которые стелются по ее поверхности. Часто уже к этому времени
происходит отмирание стержневого корня и обособление скелетных осей растения.
Несмотря на то что базальные части растений отмирают, ярко выраженных признаков
старения у обособившихся частей побегов нет. Нарастающие скелетные оси
по-прежнему активно ветвятся, обильно цветут и плодоносят, в разных частях их
побегов образуется большое количество ПД. В таком состоянии д. цельнолистная
может существовать длительное время, поэтому выявить ее предельный возраст
практически невозможно, отдельные же системы побегов в живой части имеют
возраст 50-60 лет. Несомненно, что последняя фаза онтогенеза - жизнь в виде
клона - гораздо более длительна, чем предыдущие три. Растение быстро проходит
начальные стадии развития и переходит в качественно новое состояние - самостоятельного
существования обособившихся систем побегов, которое позволяет ему значительно
продлить жизненный цикл в целом.
На Чукотке встречается иная форма роста д. цельнолистной. В
нивальных тундрах, на снежниках и карбонатных болотах этот вид нередко образует
крупные выпуклые подушки с мощным, сохраняющимся в течение всей жизни особи
главным корнем. Придаточное укоренение у таких экземпляров играет вспомогательную
роль [Юрцев, 1984).
Таким образом, в разных экологических условиях д.
цельнолистная может формировать биоморфы, принципиально отличающиеся друг от
друга. Первая экобиоморфа - стланичек с распростертыми по поверхности субстрата
ветвями, способными существовать самостоятельно с помощью активно отрастающих
придаточных корней. Вторая представлена подушковидной формой с компактно
располагающимися побегами и стержневым корнем, сохраняющимся до конца
онтогенеза; придаточное укоренение ослабленное.
ДРИАДА АЯНСКАЯ DRYAS AJANENSIS JUZ.
Дриада аянская - охотско-амфиберингийский океанический
арктоальпийский вид [Юрцев, 1984J. На исследуемой территории обычен в
Прибрежно-Охотском, Гижигинском, Охотско-Колымском водораздельном
флористических районах.
Материал собран в окрестностях Магадана на п-ове Старицкого
на щебнистой платообразной вершине сопки, расположенной на высоте 500 м над ур.
м. Плато редко заселено растительностью. Вместе с д. аянской здесь встречаются
небольшими пятнами (от 30 до 100 см) толокнянка альпийская, кассиопа
вересковидная, мытник охотский.
В онтогенезе выделено 4 фазы: 1) проросток (1-й год жизни),
2) формирование СПП (1-8-й год), 3) вычленение основных СО (8-20-й годы), 4)
жизнь в виде клона (с 20-23 лет).
П е р в а я ф а з а. Проросток аналогичен проростку
предыдущего вида.
В т о р а я ф а з а. Нарастание ПП продолжается 2-3 года,
после чего происходит первое ветвление. В результате образуется 5-6 ПВ. Часть
их отмирает в течение 1-3 последующих лет, остальные 3 (чаще 1-2) продолжают рост.
Сам ПП растет еще 2-3 года и ветвится вторично, давая уже до 8 ПВ, из которых
также 1-2 ПВ -становятся многолетними. На 8-й год (иногда раньше) ПП дает
репродуктивный прирост. После цветения происходит замещение ПП побегами
ветвления, образовавшимися под ним. ПВ, как и ПП, ветвятся через 1-2 года. Эта
цикличность сохраняется на протяжении всего онтогенеза. Определенная часть ПВ
отмирает в течение примерно 5 лет. ПВ высших порядков имеют большие размеры
приростов (1-1,5 см) по сравнению с размерами у низших порядков (5 мм). Все
побеги полегают уже к концу первого вегетационного периода, над субстратом
возвышаются верхушки побегов (5-8 мм).
К 8-летнему возрасту приросты первых 3 лет оказываются в
почве, остальная часть побегов лежит на ее поверхности, однако частички почвы и
опад прикрывают побеги, формируя плотное образование, воспринимаемое как
субстрат. Придаточные корни развиваются уже на приростах 3-4 последних лет. С
увеличением возраста некоторые из них находящиеся в основании СО, могут
значительно усиливаться, почти достигая размера стержневого корня. На 6-7-й год
появляются первые ПД. Растут они 2-3 года, чаще бывают эфемерными.
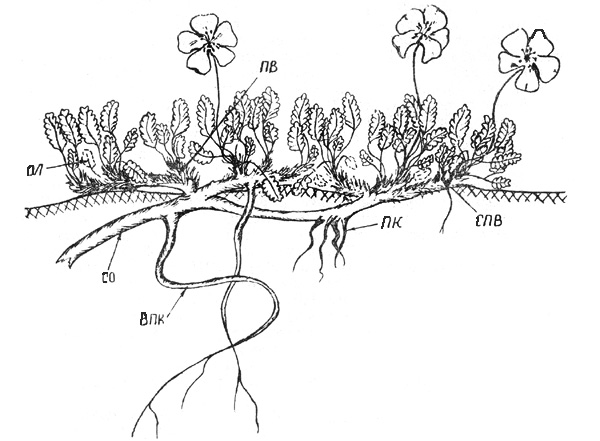
Рис. 39. Дриада аянская. Часть основной скелетной оси. ВПК -
вторичностержневые ПК
|
Т р е т ь я ф а з а. С развитием репродуктивных побегов
моноподиальное ветвление побегов сменяется симподиальным. Под цветоносом
образуется 4-9 замещающих ПВ. Один из них, как правило, растет по направлению
роста материнского (который расположен ближе всех к его верхушке), часть оставшихся
отмирает, другая часть (1-4) растет под углом к материнской оси. На следующий
год замещающие ПВ могут давать вегетативный или репродуктивный побеги. В
течение нескольких лет цикл развития побегов не меняется. Так как величина
годичных приростов невелика. а число листьев в генерации достаточно большое,
побеги располагаются скученно, образуя плотную куртину, по структуре
напоминающую подушку. Однако бывает, что некоторые из замещающих ПВ дают
удлиненные приросты (подобные ПФ). Обычно это происходит в том случае, если
нарастание продолжают 1 или 2 побега, а остальные отмирают. Длина такого побега
3 см, на нем уже в следующем году образуется большое количество ПВ и ПД.
Плотность куртины сохраняется, но подушковидность утрачивается, так как
удлиненные побеги быстрее, чем обычные удаляются от стержневого корня. В этом
случае образуется уплощенная латка с распростертыми по субстрату ветвями.
Ч е т в е р т а я ф а з а. Придаточные корни у д. аянской
обильны, но не все равноценны. Одни из них имеют небольшие размеры (от 2 до 5
см), тонкие, иногда волосовидные. Другие, напротив, лишь немного уступают
стержневому корню (рис. 39).
У 15-17 -летнего растения толщина стержневого корня у
основания 1,5 см, длина 40 см. Некоторые придаточные корни у основания толщиной
до 0,8-0,9 см, длиной от 18 до 32 см. Стержневидные корни растут сначала
геотропно, затем изгибаются и растут параллельно поверхности почвы. Мелкие
придаточные корни обычно растут по всей длине СО, иногда располагаясь пучком.
Чаще они образуются в местах ветвления побегов.
Разрушение куста происходит в месте отхождения СО от
стержневого корня или в месте образования стержневидных придаточных корней,
когда растение достигает 20-25-летнего возраста. Ветви клона продолжают
существовать отдельно неопределенно долгое время. Часто такую отошедшую ветвь
можно принять за самостоятельный куст, так как она имеет крупный, сходный со
стержневым, корень. Однако растущие побеги такого растения направлены в одну
сторону от корня, тогда как в материнском кусте СО располагаются, как правило,
радиально.
Наличие у дриады аянской стержневидных придаточных корней
может быть использовано как альтернативный признак при сопоставлении близких в
систематическом отношении видов, какими являются, например, дриады точечная и
аянская, плохо различимые по признакам, принятым у систематиков, Дриада
точечная не образует корней, подобных стержневидным корням дриады аянской. |
ТРАНСФОРМАЦИЯ ФОРМ РОСТА В РОДЕ DRYAS L.
Дриады И. Г. Серебряков [1962] относит к
вегетативно-подвижным кустарничкам шпалерного типа. Однако анализируя
морфогенез дриады восьмилепестной, он говорит о том, что у нее много черт,
присущих травянистым видам. Это позволяет ему сделать вывод, что дриада
восьмилепестная - скорее всего, переходящий тип от кустарниковых к травянистым
формам.
Процессы, приводящие к образованию трав, разнообразны и в
достаточной степени проявляются уже у древесных форм. Эта мысль неоднократно
высказывалась исследователями жизненных форм растений [Гатцук и др., 1971,
1974; Серебрякова, 1972, 1973; Хохряков, 1975, 1976б; и др.]. Чтобы судить о
трансформации древесных форм в травянистые, необходимо руководствоваться
определенными критериями, которые свидетельствовали бы о «накоплении черт
травянистости» у тех или иных форм. Существует множество различных подходов для
выявления сущности понятия «травы», однако до сего времени данный вопрос в
литературе остается дискуссионным. Подробный анализ существующих направлений по
данному вопросу дается в работе Л. Е. Гатцук [1976]. Как справедливо отмечает
А. п. Хохряков [1976БJ, безуспешность попыток дать исчерпывающее определение
этому понятию свидетельствует, очевидно, о том, что во многих линиях развития -
«филах» - уровни травянистости не совпадают друг с другом и выражаются
качественно различно.
На основе сравнительного анализа онтогенезов 4 видов дриад и
их экобиоморф выстраивается морфологический ряд: дриада большая - д. точечная -
д. цельнолистная - д. аянская, в котором прослеживаются изменения в строении
побегового тела растения, накопление «черт травянистости» от первого к
четвертому члену ряда. В качестве основного критерия «травянистости» берется
интенсивность образования придаточной корневой системы. Обоснование данного
критерия как одного из важнейших у стланиковых форм приводится в работе А. П.
Хохрякова [1976б). Анализируются и некоторые другие признаки (см. таблицу).
Изменение форм роста прослеживается уже у экобиоморф отдельных видов.
Основные параметры дриад Северо-Востока СССР
| Вид |
|
Возраст, лет |
Количество дочерних побегов |
Величина орто-
тропной части, см |
| полегания ПП |
образования ПК |
отмирания СК |
ветвления |
замещения |
| Дриада большая |
1
2
3 |
4
4
3 |
10-12
10-12
8-10 |
не отмирает
40-50
30-40 |
3-5
5-6
6-8 |
3-5
3-6
4-7 |
8
4-5
3-4 |
| Дриада точечная |
3
2 |
|
|
|
|
|
|
| Дриада цельнолистная |
1 |
1 |
4-5 |
25-30 |
6-10 |
6-10 |
2-3 |
| Дриада аянская |
1 |
1 |
4-5 |
23-25 |
5-8 |
4-9 |
0,5-0,8 |
Сравнивая онтогенез трех экобиоморф д. большой, видим, что
растение оптимальных местообитаний - настоящий кустарничек с побегами,
приподнимающимися над субстратом до 8 см, имеющий хорошо развитый стержневой
корень, который сохраняется до конца онтогенеза, с поздним развитием
придаточных корней. Годичные приросты размером 4-5 мм, листья до 5,5 см. В
экстремальных условиях это стланичек, величина ортотропных побегов которого 3-4
см, годичных приростов 1,5-2 мм, листьев 2,5-3 см. Стержневой корень отмирает в
30-40 лет, сменяясь системой придаточных корней, которые появляются уже на 8-10-летних
побегах. Позже образуется клон из обособившихся СО, которые продолжают
существовать самостоятельно. В экстремальных условиях экобиоморфа д. большой приближается по величине
годичных приростов, приподнимающихся частей побегов к д. точечной, растущей в
благоприятных условиях. Однако те особи д. точечной, которые поселяются в более
суровой обстановке, по перечисленным признакам напоминают биоморфы дриад
цельнолистной и аянской. У них наблюдается геофитизация побегов до почти
полного их погружения, усиление придаточного корнеобразования и более раннее
отмирание стержневого корня.
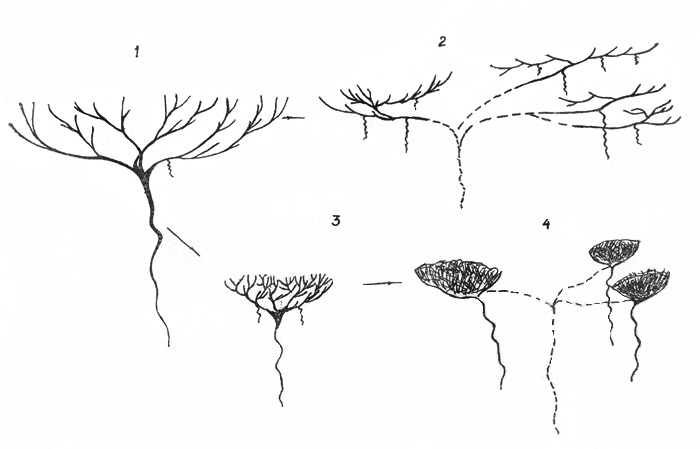
Рис . 40. Схема трансформации форм
роста у дриад: 1 – кустарничек, 2 – вегетативно-подвижный стланичек, 3 –
подушка, 4 – клон из подушковидных особей
|
В морфологическом ряду экобиоморф происходят следующие
изменения. Все более раннее а) полегание ПП, б) образование придаточной
корневой системы, в) отмирание стержневого корня и уменьшение величины
ортотропной части побегов (см. таблицу). Кроме того, всем дриадам свойственны такие адаптивные признаки, как сохранение на СО и побегах
отмерших листьев в течение длительного времени, погружение СО в грунт, причем
последним членам ряда (д. цельнолистная, д. аянская) эти признаки свойственны в
большей мере, чем первым.
Таким образом, можно говорить об усилении «черт
травянистости» у биоморф дриад, стоящих, последними в морфологическом ряду, т.
е. о тенденции к преобразованию древесных форм в травянистые.
Другое направление трансформации форм роста дриад приводит к
образованию подушковидной формы. Уменьшение величины годичных побегов,
междоузлий и одновременно интенсификация ветвления и увеличение числа
возобновляющихся побегов, которые и после отмирания остаются в куртине, а также
сохраняющиеся на побеге отмершие листья, черешки и застревающие среди них
частички почвы, дресвы, слоевищ лишайников и прочее, способствуют увеличению
компактности куста, что в конечном итоге ведет к формированию подушки. Такие
подушковидные растения отмечены для д. цельнолистной [Юрцев, 1984]. Подобные
формы образует и д. аянская. Отличие подушек этих 2 видов дриад в том, что у первых до конца онтогенеза сохраняется мощный
стержневой корень, вторые образуют придаточные корни, которые усиливаются до
размера стержневого. Наличие их обусловило способность биоморфы к
клонообразованию. Каждый парциальный куст клона обладает всеми чертами
подушковидного растения. В надземной сфере это компактность, уплотненность
побегов, в подземной - вторично-стержневой корень, аналогичный по структуре
собственно стержневому (рис. 40). |
АКОМАСТИЛИС ЛЕДЯНОЙ ACOMASТILIS GLACIALIS (ADAM.) KHOKHR.
Акомастилис ледяной - голарктический вид. Ареал охватывает
арктическую Сибирь, Чукотку, Западную Сибирь (Приобье), Восточную Сибирь
(водораздел рек Колыма и Лена), Даурию, Камчатку, бассейн р. Анадырь, а также
территорию Северной Америки [Юзепчук, 1941J. В пределах изучаемого региона
распространен во всех флористических районах, исключая Прибрежно-Охотский и
Гижигинский.
Акомастилис неоднократно привлекал к себе внимание
исследователей. Различным аспектам биологии посвящена работа Б. А. Тихомирова и
Г. И. Галазия [1952], в которой разработана методика определения возраста и
продолжительности жизни, определен температурный режим различных частей
растения. В другой работе Б. А. Тихомирова [l963] освещены вопросы
биологического значения опушения плодов, а также способность к вегетативному
размножению с помощью укороченных корневищ, затрагиваются вопросы перехода
древесных форм роста в травянистые. По мнению Б. А. Тихомирова, в условиях
Арктики переход от древесной формы роста к травянистой осуществляется через
своеобразные биоморфы, сходные с полукустарничками аридных областей, типа Sieversia
glacialis (Adam.) R. Вг. (=Acomastilis
glacialis (Adam.) Khokhr.), Potentilla elegans Cham. et Schlecht.
Материал собран в нескольких пунктах Магаданской области в
течение полевых сезонов 1978, 1980 и 1981 гг. в Сусуманском (устье р.
Урультун), Северо-Эвенском (верховье р. Омолон), Ольском (верховье р. Буюнда),
Хасынском (верховье р. Ола), Тенькинском (верховье р. Детрин) районах.
Ниже дается описание онтогенеза акомастилиса в южных
пределах его ареала: Хасынском районе (верховье р. Ола - Ольское базальтовое
плато). Популяция занимает площадь 20 м2 среди каменистых россыпей и щебенки на
платообразной вершине горы Ледяная (660 м над ур. м.). В пределах юга
Колымского нагорья более предпочтительными местообитаниями являются
дренированные, хорошо увлажненные, достаточно густо задернованные приручьевые
склоны. Однако на большей части территории Северо-Востока условия его
произрастания более близки к условиям Ольского плато и являются для него
типичными.
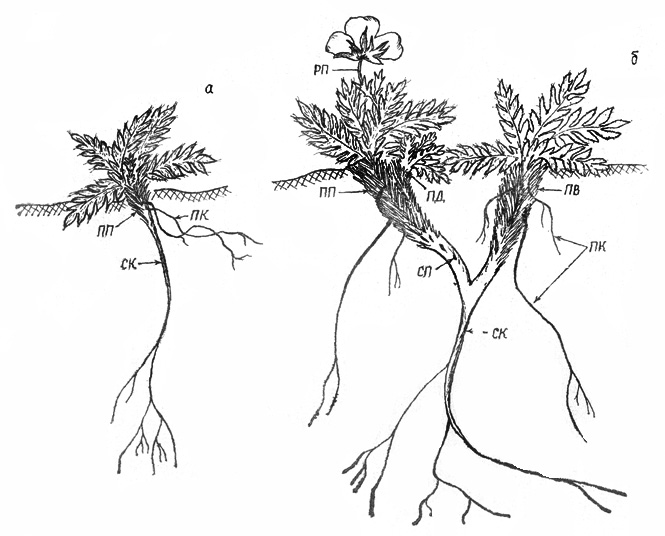
Рис. 41. Акомастилис ледяной: а – 6-й год (первая фаза), б –
ветвление ПП (вторая фаза)
|
Акомастилис в онтогенезе проходит 4 фазы: 1) нарастание ПП,
2) развитие системы ПП, 3) образование куртины, 4) образование клона. Переход
от одной фазы к другой не отграничен четко по годам, например, вторая фаза,
начинающаяся с момента ветвления ПП, у разных экземпляров приходится на б-й,
10-15-й или даже на 25-30-й годы. То же можно сказать о четвертой фазе, которая
может начинаться в 35-40 или около 100 лет, поэтому при выделении фаз
онтогенеза их длительность не указана, но обсуждается в тексте при описании
каждой фазы в отдельности.
П е р в а я ф а з а. Проростки встречаются исключительно
редко, на мелкоземистых пятнах. Прорастание надземное. К концу вегетационного
периода размер проростка 3-5 мм, с овальными семядолями и 1-2 листочками
размером до 1 см. Листовая пластинка гофрированная, слабо рассеченная по краю.
Корень неразветвленный, длиной 5-7 см. В течение нескольких лет ПП нарастает,
годичные приросты по 1-1,5 мм. Размер листьев постепенно увеличивается и к 5
годам вместе с черешками составляет 2-3 см. Черешки переходят в сильно
расширенные основания, которые полностью охватывают стебель. Форма листьев
усложняется и к 5 годам становится как у взрослых растений -
непарно-перистосложной, рассеченной до средней жилки. Листья, черешки, молодые
стебли опушены довольно длинными волосками 1,5-2,5 мм. ПП розеточной структуры,
узлы сближены, в розетке 4-5 листьев, причем пятый к концу вегетационного периода
полностью не развернут. Отмершие листья остаются на побеге, со временем
обламываются, однако их основания сохраняются долго, плотно окутывая стебель,
предохраняя его и почки возобновления от воздействий низких температур. |
Вследствие перемещения ветром мелкой дресвы и щебня
основания ПП засыпаются и в погребенной части на 4-й год образуются придаточные
корни (рис. 41, а), которые выполняют двоякую функцию: обеспечивают растение
питанием и, притягивая ПП к субстрату, способствуют его постепенному
погружению. В пазухах листьев формируются почки, часть которых развертывается в
ПВ, часть других, после покоя, - в ПД, остальные (их большинство) длительное
время остаются на побеге, погружаясь вместе с ним в почву. Когда основания
черешков отгнивают и разрушаются, то на оголившихся частях стебля видны
многочисленные почки, которые часто отмирают вместе с ним (рис. 41, б, 42).
Такой погружающийся стебель, образованный укороченными годичными побегами,
морфологически соответствует эпигеогенному корневищу многолетних травянистых
растений.
В т о р а я фаз а. Первое ветвление ПП происходит иногда уже
на 5-й год, бывает на 10-15-й год или даже в 25-30 лет. Ветвление
моноподиальное, ПВ образуются 'из пазушных почек одновременно с нарастающим ПП.
При первичном ветвлении обычно возникает 1 ПВ, который так же, как и ПП,
длительно нарастает, образуя короткие годичные приросты (1-1,5 мм). К 15 годам
они и у ПП, и у ПВ становятся длиннее (2-2,5 мм). На каждом развивается розетка
с 5-7 листьями, самый крупный из них достигает 7 см. ПП, выполняющий теперь
также и функцию СО, не только постепенно удлиняется, но и увеличивается в
диаметре. У взрослых многолетних экземпляров диаметр погруженной многолетней
оголенной СО около 1 см (чаще 5-8 мм), и он более или менее одинаков по всей
длине оси. Ее же диаметр с сидящими остатками листьев 4-6 см. У молодых
растений он несколько меньше - 2-3 см. Диаметр надземной розетки у многолетних
экземпляров достигает 10 см и более. В розетке накапливаются частички почвы и
отмершие части растений, она уплотняется и по форме напоминает подушковидные
растения. |
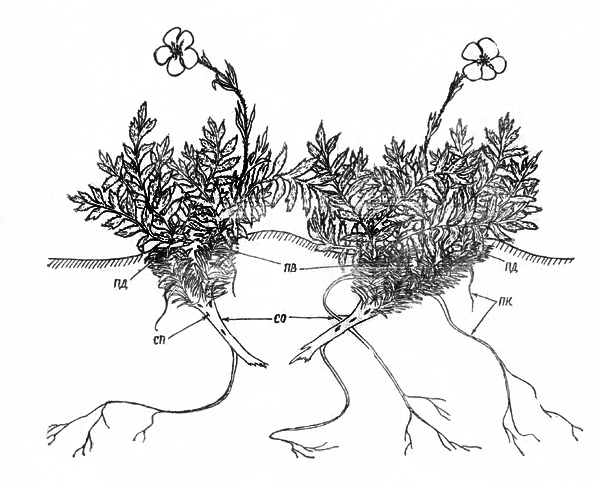
Рис. 42. Акомастилис ледяной. Фаза клона |
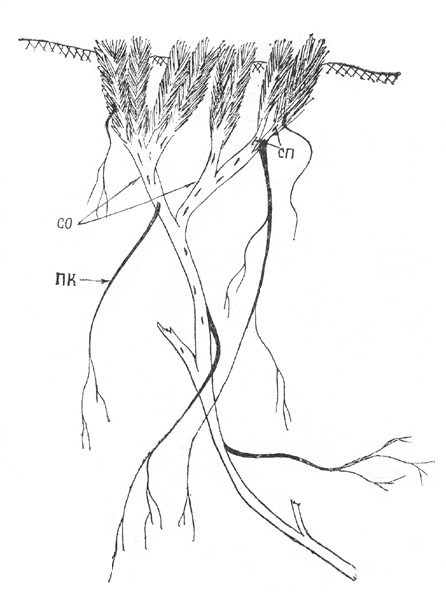
Рис. 43. Акомастилис ледяной. Компактная куртина
|
К 15 годам происходит вторичное ветвление побегов у
экземпляров, первое ветвление которых было в 5 лет. Если же оно имело место в
более позднее время - соответственно ПП ветвится вторично позже, но, как
правило, это происходит спустя 8-10 лет. У отдельных индивидуумов нарастание ПП
и 1-2 ПВ может идти в течение всей жизни до отмирания. На таких долго
нарастающих СО могут формироваться лишь быстроотмирающие и эфемерные ПR и ПД.
К 15-20 годам по-прежнему сохраняется стержневой корень, но
образование придаточных корней становится более активным, они обычны уже на
приростах 4-5 последних лет. Придаточные корни двух типов. Одни достаточно
большие (20-30 см), многолетние, другие - тонкие, волосовидные,
быстроотмирающие, их особенно много на 4-8-летних приростах СО. Над
поверхностью почвы остаются годичные побеги 4-5 последних лет, т. е. менее 1 см
длины стебля, остальная его часть вертикально заглублена в этом возрасте (15
лет) на 2,5 см.
Т р е т ь я ф а з а. С 20-30-летнего возраста или несколько
позже ПВ возникают более или менее регулярно через 5-7 лет как на ПП, так и на
ПВ первого порядка, начинается формирование СПВ. ПВ развиваются из пазушных
почек и растут под углом к материнскому побегу. Расположение их может быть
разным. На одних особях ПВ плотно прижаты друг к другу, образуя компактную
куртину, по структуре близкую к подушковидным формам. В этом случае большинство
образовавшихся ПВ в течение нескольких лет нарастает, затем ослабленные побеги
отмирают, но не обламываются, а остаются в куртине, способствуя еще большему ее
уплотнению и выполняя роль «утеплителя» для оставшихся живых побегов. Как
правило, в такой СПВ до конца онтогенеза насчитывается не более трех-четырех
порядков побегов ветвления (рис. 43). У других экземпляров ПВ отходят от СО
почти под прямым углом. После многолетнего нарастания таких ПВ СПВ оказываются
удаленными друг от друга и от СО на разные расстояния. Образуется рыхлая
куртина, занимающая площадь от 0,5 до 1 м2.
Она состоит из нескольких разных по размеру и возрасту СПВ.
Одни из составляющих куртину СПВ в своей надземной части имеют диаметр 25-30
см, у других, представленных одним неветвящимся ПВ, размер розетки 8-12 см в
поперечнике (рис. 44).
Стержневой корень к 30-40 годам отмирает. Отдельные СПВ
куртины объединены внизу одним корневищем (погруженные СО), которое заглублено
на 18-20 см. Молодые его части располагаются вертикально, старые - косо по
отношению к поверхности почвы. На старых частях корневищ остатки листьев
постепенно разлагаются, но их еще можно обнаружить на 30-40-летних годичных приростах,
хотя при выкапывании они свободно отделяются от стебля. К молодым же частям
побегов основания листьев прикреплены так крепко, что удалить их, не повредив
стебля, невозможно. |
Зацветает акомастилис в 15-18 лет. Репродуктивные побеги
развиваются из пазушных почек предыдущего года, они неразветвленные, несут 3-7
листьев, уменьшающихся к верхушке побега. Цветонос и листья обильно опушены.
Цветок 1, диаметр венчика 2,5-3,5 см. Высота цветоноса 4-5, к моменту
образования семян 8-14 см.
Как на ПП, так и на ПВ в надземной и подземной, но близкой к
поверхности частях достаточно часто появляются ПД из 3-8-летних почек. Одни из
них эфемерные, другие нарастают 3-6 лет, их годичные приросты составляют 5-7
мм. ПД, развивающиеся из подземных почек, в 1-й же год пробиваются на
поверхность и образуют розетку с 5-7 листьями. Такие ПД на 2-3-й год могут
формировать репродуктивный побег, но после цветения отмирают полностью. К 30
годам годичные приросты ПП и ПВ увеличиваются до 3-4 мм. Над поверхностью почвы
растения возвышаются на 2-3 см. В розетке развивается 13-15 листьев, на каждый
миллиметр длины стебля приходится 3-4 листа.
Ч е т в е р т а я ф а з а. После отмирания стержневого
корня начинается постепенное разрушение базальных частей корневища. Функцию
питания выполняют придаточные корни, которые достаточно равномерно распределены
по СО. На старых частях сохраняются вторичностержневые корни, на молодых, кроме
них, - множество мелких быстроотмирающих корней. Часто, но не всегда, отдельные
СО (многолетние СПВ) теряют связь друг с другом вследствие разрушения их
базальных частей, после чего они существуют самостоятельно. Распадение куртины
обычно происходит в возрасте около 100 лет, но бывает и значительно раньше - в
45-50 лет; образуется клон. Границы его, как правило, хорошо определимы, так
как возникающие партикулы расположены достаточно близко друг к другу далеко от
других клонов. |
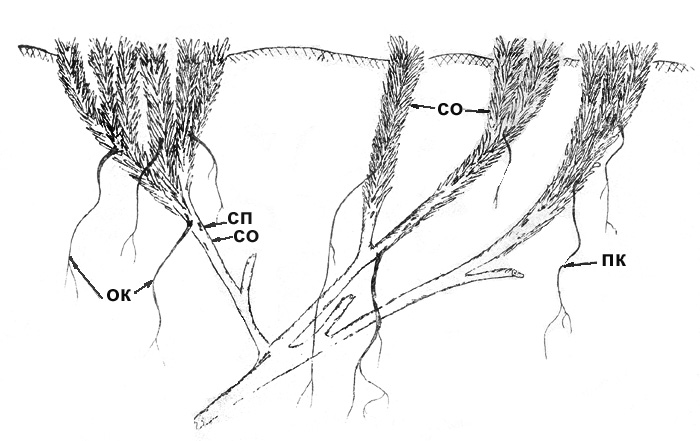
Рис. 44. Акомастилис ледяной. Разросшаяся куртина |
Возраст клона установить невозможно, так как нельзя
вычислить возраст его отгнившей части. Полностью отмерший клон в популяции не
встречен, однако стареющие экземпляры с отдельными отмирающими или отмершими
партикулами есть. Предельный возраст сохранившейся части таких экземпляров
определен в 120-130 лет. В деградирующих парциальных СО системы побегов
начинают отмирать с более слабых ПВ и их систем. У отдельных СПВ последней
отмирает СО, несущая на себе всю систему ПВ. Многолетние СО в конце онтогенеза
образуют только эфемерные ПВ и ПД, но и они со временем перестают появляться и
СО отмирает. Сенильные растения в популяции встречаются редко, в основном
популяция представлена молодыми и средневозрастными.
Наблюдения за растениями в более благоприятных условиях
показали, что нарастание систем побегов в онтогенезе происходит аналогично описанному
выше, однако размеры растений и их отдельных частей могут значительно
варьировать. На задернованном увлажненном склоне в устье р. Урультун куртины
возвышаются над уровнем почвы до 6, иногда до 8 см. Размер листьев 7-8 см. СО
располагаются параллельно поверхности почвы на глубине 10-15 см, наибольшая
длина СО у одного из самых крупных экземпляров 49 см. От основания эта СО была
разрушена, приблизительный возраст сохранившейся части 140-150 лет.
В течение всего онтогенеза у акомастилиса происходит нарастание
системы ПП, сохраняется моноподиальное ветвление. Базальные части стебля
погружаются в почву, формируя эпигеогенное корневище. Вследствие последующего
разрушения стареющих частей корневища к концу онтогенеза прослеживается
тенденция к вегетативной подвижности, однако это нельзя считать полным,
переходом к активному передвижению по площади. Продвинутость такой системы в
сторону активизации вегетативной подвижности несомненна, о чем свидетельствует
и наличие усиленного формирования придаточной корневой системы. СО в надземной
сфере развивают систему побегов, по структуре близкую подушковидной форме
роста. В конце жизненного цикла образуется клон с несколькими самостоятельно
существующими СО.
Своеобразие жизненной формы акомастилиса в том, что она может
быть представлена и как розеточный короткокорневищный травянистый многолетник,
и как погруженная эпигеогенно-корневищная подушка.
СИВЕРСИЯ МАЛАЯ SIEVERSIA PUSILLA (GAERTN.) HULT.
Сиверсия малая - североохотско-корякский арктоальпийский
(преимущественно высокогорный) субокеанический вид [Юрцев, 1984J. В исследуемом
регионе массовое растение от Охотского до Колымского флористических районов,
реже встречается в Омолонско-Анюйском, Анадырском, Корякском районах. Распространена
в кустарниковых тундрах, на альпийских лужайках, в зарослях кедрового стланика.
Здесь приводится описание онтогенеза сиверсии малой по
материалам, собранным в 1976 г. в Тенькинском районе Магаданской области близ
пос. Кулу. Растение можно обнаружить на горных тундровых участках на высоте
850-900 м над ур. м. Сиверсия тяготеет к влажным местообитаниям - поселяется
либо вблизи снежников, либо в увлажненной разнотравно-кустарничковой тундре и,
как правило, на мелкощебнистом субстрате.
В онтогенезе выделено 4 фазы: 1) нарастание ПП (1-4-й годы),
2) развитие системы ПП (2-7-й годы), 3) формирование куртины (8-15-й годы), 4)
жизнь в виде клона (с 16-20 лет).
П е р в а я ф а з а. Проростки можно найти на голых пятнах
щебенки рядом со взрослыми растениями. Но так как сиверсия - обитатель влажных
тундровых участков, которые плотно заселены растительностью, то встречаются они
довольно редко.
К концу 1-го года вегетации проростки представляют собой
укороченный, не более 1 см, ортотропный побег розеточного типа с овальными
пожелтевшими семядолями и 5-7 листочками. Первые листья очень мелкие (длина
центральной доли 1,-2 мм), слаборассеченные.
К исходу августа листья желтеют, но не опадают, а остаются
на стебле. Стержневой корень 3-5 см (рис. 45, а). в таком виде растение уходит
под снег. На следующий год после стаивания снега и прогрева почвы терминальная
почка ПП трогается в рост и дает прирост в 2-3 мм, на котором развертывается
5-7 листьев размером чуть крупнее, чем в предыдущем году. На 2-4-й год
происходит незначительное увеличение ПП в размерах. К концу четвертого
вегетационного периода ПП 1,5 см, длина корня 13-15 см.
В т о р а я ф а з а. Ветвление у сиверсии моноподиального
типа начинается на 2-й, чаще на 4-й год развития ПП, Почки заложенные в пазухах
листьев предыдущего года, раскрываются одновременно с верхушечной почкой ПП и
дают 1-4 ПВ. Последние растут косо вверх, ослабленные и в большинстве случаев
отмирают к концу вегетационного периода: Лишь 1 (очень редко 2) наиболее
сильный ПВ сохраняет способность к дальнейшему росту. Приросты ПВ не превышают
приростов ПП. Размер листьев 5-6 мм, черешков - 6-7 мм. В розетке 8-9 листьев.
Нарастание ПП и ПВ продолжается не более 3 лет, после чего они отмирают.
Развитие сиверсии на данном этапе не всегда соответствует
описанной схеме. В отдельные годы все образовавшиеся ПВ отмирают и происходит
либо вторичное ветвление ПП, либо в основании его на первом приросте развиваются
из почек, покоившихся в течение 2-3 лет, ПД. Эти побеги обычно эфемерные и
отмирают в этом же году, но иногда растут 2-3 года. Корень 6-7-летнего растения
увеличивается до 18 см (рис. 45, б). Предельный возраст ПП не более 7 лет.
Однако на 4-5-й год в базальной его части, скрытой в почве, из почек прироста
1-го года развиваются столоновидные побеги (гипогеогенные корневища). |
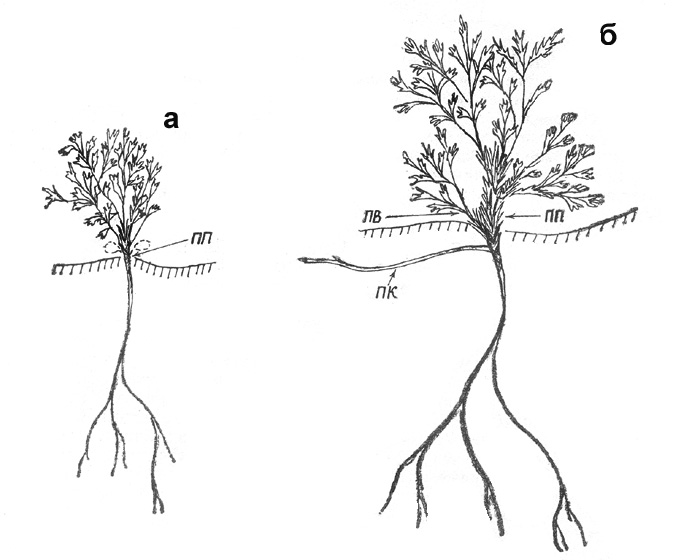
Рис. 45. Сиверсия малая; а - проросток, б - начало ветвления
и отрастания корневищ |
Т р е т ь я ф а з а. Столоновидные побеги (по
классификации М. Т. Мазуренко и А. П. Хохрякова [1977], ПФ) отличаются от всех
ранее описанных побегов многими признаками. Во-первых, они образуются из почек,
покоившихся в течение 2-5 лет. Во-вторых, они плагиотропные и обладают
усиленным ростом (величина годичных приростов варьирует от 2 до 10 см и более).
В-третьих, они не сразу выходят на поверхность, а в течение 5 лет и более
растут в почве на глубине 2-4 см. В-четвертых, основная функция всех надземных
побегов, как известно, - фотосинтез и плодоношение, функция подземных
столоновидных побегов - завоевание площади, формирование куртины. Побеги
столоновидного типа были описаны ранее у кустарничков Т. Г. Дервиз-Соколовой
[1966] на примере некоторых простратных форм ив. М. Т. Мазуренко [1978]
отмечает наличие их у кустарничков гольцов ого и подгольцового поясов:
толокнянки альпийской, рододендрона камчатского, брусники, голубики и некоторых
других. На 4-7-й год в основании ПП трогаются в рост 2-4 почки.
Побеги вначале растут горизонтально в почве, но после отмирания надземной части
ПП (или еще до этого) 2-3 из них выходят на поверхность. Длина плагиотропной
надземной части таких побегов 2-3 см, затем верхушка их приподнимается и
развивает розетку из 3-5 листьев.
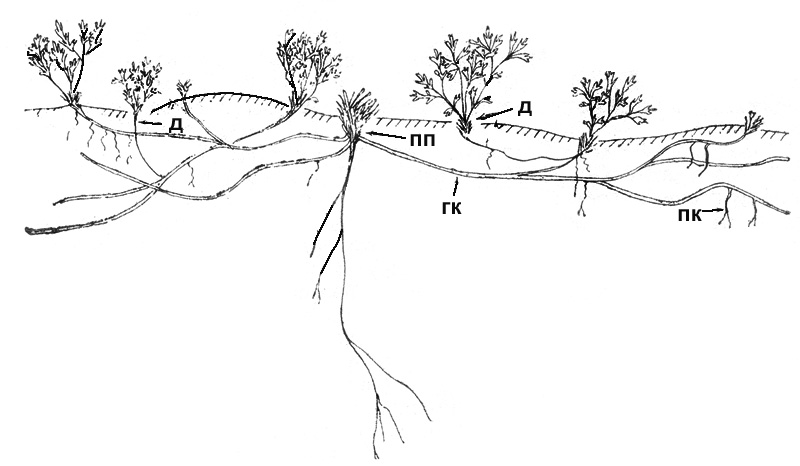
Рис. 46. Сиверсия малая. Разросшаяся куртина
|
Дальше идет формирование дочерних кустов. Приросты побегов
3-5 мм, они ветвятся, живут от 3 до 5 лет. В грунте столоновидные побеги также
активно ветвятся, развивая многочисленные ПВ, которые после 3-4 лет дают ПВ
следующего порядка и т. д. Отдельные ПВ выходят на поверхность и формируют
дочерний куст, в котором уже через 1 год появляются цветоносы, а в подземной
части - новые столоновидные побеги. В разных частях побегов образуются ПД. Если
они развиваются на подземных частях, то в год своего образования усиленно
растут (прирост 3-4 см), некоторое время стелются по поверхности, потом дают
ортотропный прирост. Иногда они развивают цветонос, но после цветения отмирают.
В надземной части растения ПД появляются в основании ортотропных частей
побегов, они ослабленные и отмирают в конце первого вегетационного периода.
Репродуктивный прирост (цветонос) развивается из
терминальной почки ортотропного побега. После цветения последний отмирает, но в
основании его из подземных почек отрастают 2-3 замещающих побега ветвления.
Замещающие побеги имеют восходяще-ортотропное направление роста, они в 1-й же
год выходят на поверхность и к следующему году формируют цветочные почки.
Иногда цветок появляется на 2-й или 3-й год развития побега, т. е. цветоносные
побеги сивер сии могут быть моно-, ди- и трициклическими. Несмотря на то что
побеги, отходящие от стержневого корня, долго сохраняют с ним связь, для
сиверсии характерно раннее появление придаточных корней. Довольно много их
образуется в основании ортотропных частей, а на подземных побегах они
появляются уже на приросте предыдущего года (рис. 46). |
Сиверсия образует множество надземных и подземных побегов,
формируя куртину, диаметр которой к 16-20 годам составляет 60-80 см.
Репродуктивный прирост несет крупный (до 2 см в диаметре) белый цветок, к
моменту плодоношения цветоножка которого вытягивается и достигает длины 6-7 см.
Листья до 1,5 см длиной, растут мутовкой, длина черешков 2,5 см. Розетковидный
надземный побег возвышается над субстратом до 3,5-4 см. Листья к концу
вегетационного периода засыхают, черешки обламываются, но их основания плотно
укрывают побег. Отдельные дочерние кустики так далеко отстоят от материнского,
что определить их принадлежность к данной куртине не всегда возможно. Побеги
тонкие (не более 3 мм в диаметре), поэтому выкопать растение без повреждения не
всегда удается.
Ч е т в е р т а я ф а з а. Приблизительно к 20-летнему
возрасту отмирает стержневой корень, разрушаются базальные части столоновидных
побегов. Дочерние кустики продолжают существовать самостоятельно (рис. 47).
Функции питания, прикрепления к субстрату полностью ложатся на придаточные
корни, которых особенно много, как упоминалось, в основании ортотропных
побегов. Последние являются центром образования новых систем побегов, цикл
развития которых аналогичен развитию системы ПП. Происходит партикуляция клона,
хотя отдельные парциальные особи длительное время сохраняют связь друг с другом.
Столоновидные побеги отрастают уже от оснований парциальных кустов, растут,
выносят к свету новые надземные розетки, отстоящие на значительном расстоянии
(до 18 см) от материнской особи.
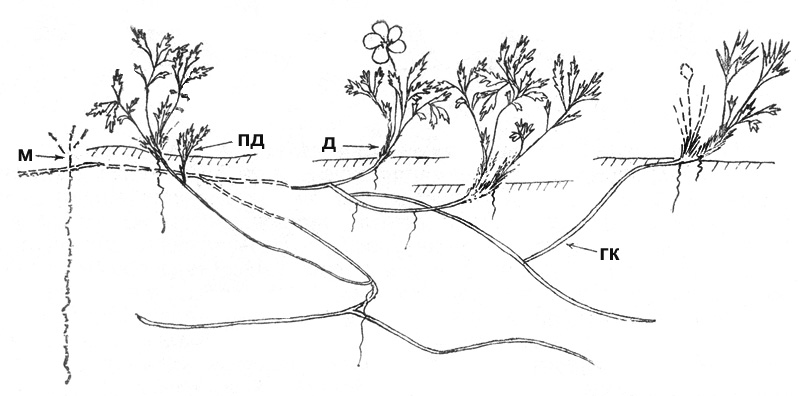
Рис. 47. Сиверсия малая. Клон
|
Все новые дочерние кусты образуются в клоне, которые, в свою
очередь, образуют подземные столоны. Происходит разрастание клона, который
может занимать большое пространство, используя любой свободный участок для
выноса надземных побегов. Многие соседствующие с сиверсией растения не выносят
конкуренции и вытесняются ею. Поэтому нередко можно встретить довольно большие
участки, полностью покрытые как ковром сиверсией малой. Растения других видов
либо угнетены, либо вообще отсутствуют.
Установить предельный возраст сиверсии не удалось. С большей
или меньшей точностью о ее возрасте можно говорить в первые фазы онтогенеза, приблизительно
до 20-25 лет. Трудность определения возраста зависит от ряда причин. Во-первых,
величина годичных приростов столоновидных побегов сильно варьирует и отличить
один годичный прирост от другого сложно. Во-вторых, с момента партикуляции и
отмирания стержневого корня невозможно определить принадлежность отдельных
особей к какому-либо клону. В-третьих, нам не удалось встретить в природе
отмирающие клоны. Однако возраст отдельных систем побегов, сохранивших связь
друг с другом, исчисляется в несколько десятков лет (до 4 и более). Жизненную
форму сиверсии можно отнести к розеточным вегетативно-подвижным травянистым
многолетникам. |
|