Вы находитесь: НАША БОТАНИЧКА — Теория — Биоморфология — Антропова Г. Л. Биоморфология розоцветных Северо-Востока СССР
ОНТОГЕНЕЗЫ БИОМОРФ ИССЛЕДОВАННЫХ ВИДОВ
ШИПОВНИК ИГЛИСТЫЙ ROSA ACICULARIS LINDL.
Шиповник иглистый - голарктический, почти циркумполярный вид.
Обычен в светло- и темнохвойных лесах Евразии и Северной Америки. На территории
Советского Союза распространен от северо-западных границ (через Урал и всю
Сибирь) до восточных (включая Сахалин и Камчатку) и северо-восточных (Чукотка)
рубежей страны [Юзепчук, 1941]. Более подробно ареал вида представлен в
«Арктической флоре СССР» [1984]. В Магаданской области отмечен во всех
флористических районах, кроме о-ва Врангеля [Хохряков, 1985].
Как ценное лекарственное сырье шиповники давно привлекают
внимание исследователей. Существует обширная литература по этому вопросу, с
которой можно ознакомиться в «Атласе ареалов и ресурсов лекарственных растений
СССР» [1976]. Работ по исследованию онтогенеза ш. иглистого в условиях Крайнего
Севера нет, есть краткое изложение жизненного цикла парциального куста
[Мазуренко, Хохряков, 1976, 1977] и краткое описание этапов органогенеза
[Назарова, 1979].
Материал собран в полевой период 1983 г. в; верховьях р.
Кулу (юго-западные отроги Колымского нагорья) и в 1989 г. на стационаре
«Контакт».
Ш. иглистый - вид с широкой экологической амплитудой, он
заселяет различные экотопы лесного пояса, речных долин н террас, поднимается на
каменистые склоны, преимущественно южные, но в гольцовый пояс не заходит. В
исследуемом регионе выделено четыре типа местообитаний вида.
Первый тип. Надпойменная терраса с древесным ярусом из
лиственницы даурской (редколесье) и кустарникового полога из ш. иглистого и
некоторых видов ив. Эдафические условия оптимальные - почвы влажные, хорошо
дренированные. Благодаря редкостойности древесного яруса растения получают
много света и одновременно защищены от воздействия ветра.
Второй тип. Невысокие (20-40 м) холмы в долине реки, которые
сложены мелко- и крупнощебнистыми породами и достаточно плотно заселены
травянистой растительностью. Склоны холмов рано освобождаются от снега, почва
быстро прогревается, поэтому растения ш. иглистого начинают раньше
вегетировать, соответственно быстрее наступают и другие фенологические фазы -
цветение и плодоношение.
Третий тип. Подгольцовая часть склонов южной. экспозиции несколько
выше границы леса. Растения не защищены от ветра, находятся в условиях
недостаточного увлажнения, глубина снежного покрова незначительна. Почвенный
слой маломощный, преобладают скелетные и фрагментарные почвы.
Четвертый тип. Затопляемые песчаные и галечные отмели в
поймах рек. Растения подвержены постоянному ветровому воздействию (эффект
«русловой трубы»), растут на слаборазвитых почвах в условиях периодического
переувлажнения - 2-3 раза за сезон отмели частично или полностью затопляются
паводковыми водами. Этим четырем типам местообитаний соответствуют четыре
экобиоморфы ш. иглистого.
Экобиоморфа 1
Верховья р. Кулу, опушка разреженного пойменного
лиственничника. Проростки и молодые экземпляры можно обнаружить в укромных
защищенных местах: под берегом, около крупных камней, но встречаются они
довольно редко.
Для всех экобиоморф ш. иглистого характерны следующие фазы
онтогенеза: 1) нарастание, ПП (1-6-й годы), 2). формирование первичного куста
(с 6 до 15-18 лет), 3) разрастание систем побегов, образование парциальных
кустов (с 15-18 до 40 лет), 4) формирование клона (с 40 лет). Время наступления
и длительность фаз онтогенеза у разных экобиоморф могут варьировать.
П е р в а я ф а з а. Семена прорастают в конце июня-начале июля.
Прорастание надземное, вслед за семядолями развертываются 2-3 настоящих листа
(рис. 2, а). ПП ортотропный. К концу вегетационного периода 5 мм, корень 4-5
см. Нарастает 3-4 года, ПП увеличивается до 3,5 см, основание его постепенно
присыпается почвой, корень - 8-10 см, слабоветвистый, с 3-4 порядками боковых
корешков.
В т о р а я ф а з а. К 5 годам ПП отмирает до уровня
почвы. От живой части его основания отрастают 1-2 ПФ длиной 2-4 см, с 2-4
листьями. На погруженной части побегов развиваются придаточные корни (рис. 2,
б). К следующему году верхушка ПФ отмирает, а приверхушечная часть симподиально
ветвится. ПВ 1 см длиной, с 2-3 листьями, чаще эфемерные, но иногда нарастающие
1-2 года не увеличивая длины прироста. Цикл развития ПФ первого поколения 3-4
года. На 8-й год появляются 1-2 ПФ второго поколения 4-5 см длиной с циклом
развития не более 3 лет. В каждый следующий год в кусте отрастают новые ПФ все
более высоких порядков, которые начинают рост еще до отмирания побегов
предыдущего порядка. Основания побегов формируют узел кущения, который со
временем разрастается, утолщается, несет большое число почек возобновления
(рис. 2, в). С возрастанием порядка ПФ постепенно увеличивается цикл их развития, сначала до
5, позже до 7 лет. |
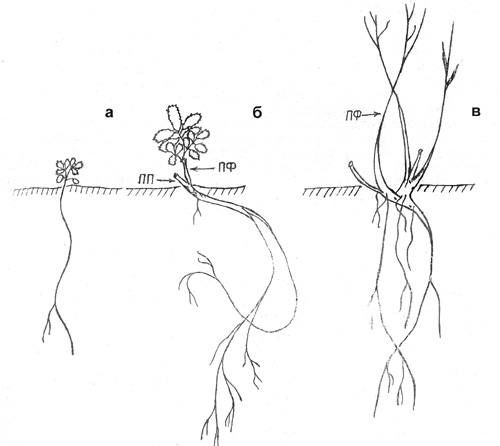
Рис. 2. Шиповник иглистый: а – проросток; б – 5-й год
онтогенеза; в – 12-13-й год онтогенеза (вторая фаза) |
Длина годичного прироста в 1-й год развития всегда бывает
наибольшей и постепенно достигает у ш. иглистого 80-100 см. На следующий год
верхушка его отмирает, а на живой части отрастает несколько ПВ от 3 до 10 см
длиной; образуется СПФ. Замещение СПФ низшего порядка СПФ более высокого
порядка происходит нерегулярно; они растут или одновременно, или же новые
появляются после отмирания старых. Как правило, куст состоит из 6-8
живых (молодых и стареющих) СПФ и 5-6 отмерших. Последние «отодвигаются» на
периферию куста, наклоняются и со временем обламываются.
Т р е т ь я ф а з а. В 15-18 лет - из спящих почек основания
узла кущения, которое погружено в почву, отрастают плагиотропные ПФ,
развивающиеся по типу гипогеогенных корневищ (ГК), несущие почки возобновления
и придаточные корни. Их годичные приросты от 8-10 до 20-40 см. Придаточные
корни на корневищах могут быть разными. Одни долго нарастающие (живут более 3
лет) ослабленные, по 10-12 см длиной или, что бывает чаще, достаточно мощные -
50 см длиной, другие - быстро отмирающие (живут 1-3 года). После 2-3-летнего
нарастания терминальная почка корневища или одна из его боковых почек дает
ортотропный надземный прирост 7-8 см длиной. Он является первым звеном в
формировании парциального куста, развитие которого может идти двумя путями.
1. На 2-й год надземный побег ветвится по моноподиальному
типу. ПВ укороченные - 2-5 мм с розеткой из 3-5 листьев, эфемерные. В следующем
году ПФ отмирает на треть длины, оставшаяся часть ветвится (ПВ 10-15 см). На
них развиваются ПВ второго порядка, опять укороченные (1 см), с розеткой
листьев. Затем СПФ отмирает. Основной цикл длится 4-5 лет.
2. На 2-й год надземный побег отмирает до уровня
подстилающей поверхности, но от его основания отрастают 1-4 ПФ, которые к
следующему году отмирают частично или до основания. В зависимости от этого
новые побеги появляются либо в зоне кущения, либо по всей длине оставшихся
живых частей побегов. В дальнейшем они ветвятся, затем происходит замещение
одной СПФ другой (рис. 3, а).
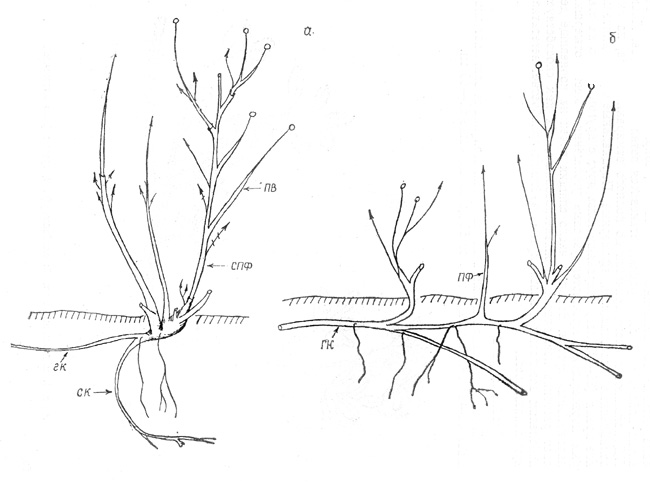
Рис. 3. Шиповник иглистый: а – первичный куст, б – часть
корневища с развивающимися парциальными кустами. Здесь и далее на рисунках: ГК
– гипогеогенное корневище, СК – стержневой корень
|
Постепенно общая высота СПФ достигает 100-110 см, а основной
цикл - 5-7. В кусте одновременно насчитывается 8-10 живых и 5-7 отмерших СПФ.
Узел кущения является зоной формирования новых надземных и подземных систем побегов.
Корневища не только нарастают в длину, но и многократно ветвятся.
Репродуктивные побеги в материнском кусте возникают на СПФ четвертого-пятого
поколения, возраст которых не менее 3 лет. Их дают терминальные почки ПВ.
Цветоносы неразветвленные с одним крупным (6 см в диаметре) цветком. В
парциальных кустах они появляются на СПФ первого поколения на 2-3-й год ее
развития.
Ч е т в е р т а я ф а з а. Общую продолжительность
онтогенеза ш. иглистого установить по ряду причин невозможно. Во-первых, в
кусте происходит постепенная смена одних СПФ другими без какой-либо последовательности.
Во-вторых, годичные приросты подземных побегов неравномерны, да и границы их не
всегда различимы. В-третьих, наиболее старые части корневищ со временем
разрушаются, парциальные кусты теряют связь друг с другом. К. тому же каждый
дочерний куст становится центром образования все новых парциальных кустов (рис.
3, б). Формируется клон из многих особей, который занимает площадь в 20-30 и
более квадратных метров. Клон состоит из разных по возрасту кустов от молодых
до стареющих и отмерших. Отмирание систем побегов в деградирующем кусте идет
следующим образом. Вначале уменьшается величина приростов ПФ, сокращается цикл
их развития вплоть до 1 года, редким становится ветвление побегов. Лишь ПД
появляются систематически до полного отмирания куста, но они ослабленные,
неветвящиеся, эфемерные. Парциальные кусты длительное время (30-35 лет)
сохраняют связь друг с другом. Часто на старых корневищах встречаются узловые
утолщения - места отмерших парциальных кустов. Большое число спящих почек на
корневище делает растение исключительно жизнеспособным, так как способствует
появлению все новых и новых кустов, растение приобретает вегетативную
подвижность. |
Экобиоморфа 2
Щебнистый склон холма в долине реки. Популяция занимает
площадь 40-50 м2. Высота кустов от 50 до 80 см, чаще 50-60 см.
Пер в а я ф а з а. Протекает аналогично предыдущей форме
роста.
В т о р а я ф а з а. Максимальный прирост 1-го года ПФ 20-25 см, кусты достаточно
густые, обычно образованы 6-8 живыми СПФ и несколькими отмершими. Отмирание ПФ 1-го
года не полное - на 2/3 или 1/2 часть длины, что вызывает более длительное
существование систем побегов растения. У экобиоморфы 2 основной цикл составляет
в среднем 9 лет.
Т р е т ь я ф а з а. К описанным ранее механизмам образования
парциальных кустов добавляется еще один. Корневище после нескольких лет
подземного роста выходит на поверхность, но к ортотропному росту не переходит,
а стелется по субстрату. ПВ, формирующиеся на нем, растут ортотропно (рис. 4).
Через год ПВ и верхушка корневища отмирают, но от живой части отрастают 1-2 ПФ
усиленного роста (10-18 см), которые и являются первым звеном в развитии
парциального куста, дальнейшее формирование которого идет по описанному выше
пути. Но бывает, что стелющееся корневище в этом же году приобретает
ортотропность и на его базе формируется новый парциальный куст.
Ч е т в е р т а я ф а з а. Разрушение подземных скелетных
осей происходит поздно, приблизительно после 50 лет. |
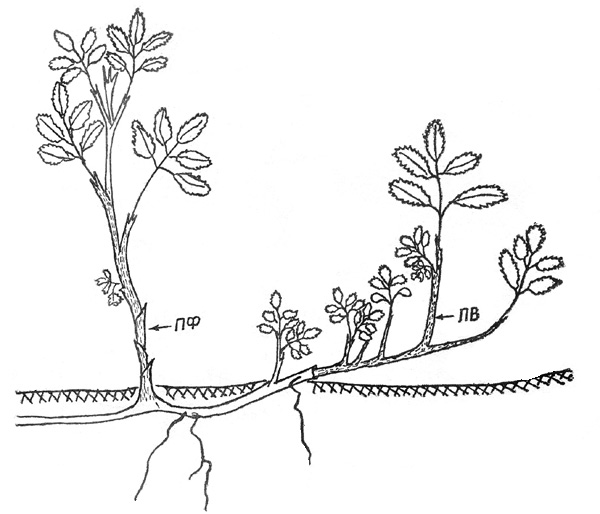
Рис. 4. Шиповник иглистый: экобиоморфа 2, третья фаза
онтогенеза |
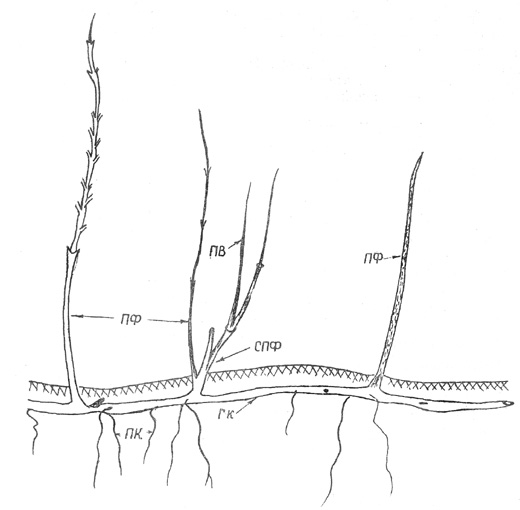
Рис. 5. Шиповник иглистый: экобиоморфа 3, часть корневища с
надземными побегами
|
Экобиоморфа 3
Подгольцовая часть склона южной экспозиции, высота над
уровнем моря 900-1000 м. Популяция занимает небольшую площадь около 10 м2.
Высота кустов 30-50 см.
П е р в а я и в т о р а я ф а з ы онтогенеза не описаны,
так как в популяции отсутствуют ювенильные формы.
Т р е т ь я ф а з а. ПФ имеют 3-, реже 4-летний цикл развития.
В 1-й год прирост ПФ составляет 25-30 см. Он неразветвленный, с листьями,
рассредоточенными по всей длине. На 2-й год верхушка ПФ отмирает, но в его
привершинной части отрастают ПВ длиной от 1,5 до 8 см. На 3-й год появляются ПВ
второго порядка, большинство их несет репродуктивные побеги. После цветения СПФ
отмирает наполовину, тогда ПВ первого порядка приобретают усиленный рост,
достигая 16-18 см. ПВ второго порядка тоже симподиальные, но меньших размеров,
чем предыдущие и несут репродуктивные органы (рис. 5).
Ч е т в е р т а я ф а з а. Популяция представлена одним
разросшимся клоном. Отдельные кусты потеряли связь друг с другом. Разрушение
корневища происходит после 25-30 лет.
Экобиоморфа 4
Песчано-галечная отмель в долине реки. Популяция занимает
площадь около 15 м2. Дважды за сезон на 3-5 дней площадь частично или полностью
покрывается водой. От предыдущих форм отличается меньшими приростами ПФ и более
сильным их отмиранием. Проростки в популяции не найдены, размножается сугубо
вегетативным путем. Репродуктивные побеги появляются на ПФ первого порядка на
2-3-й год их развития. Максимальный прирост ПФ 18-20 см, основной цикл
не более 3 лет. Образование парциальных кустов идет следующим образом. |
Вариант 1. Корневище длительное время растет в почве на
глубине 8-10 см, многократно ветвится, образуя подземную сеть. Из спящих почек
корневища развиваются надземные побеги, которые возникают без какой-либо
последовательности. Не все побеги становятся зоной формирования куста, часто
они отмирают в год своего появления, иногда нарастают до 3 лет. Как правило,
почки возобновления, находящиеся в основании побегов, погибают во время
паводков, поэтому более или менее крупные кусты образуются в том случае, если
ПФ возникают из почек, расположенных на подземных побегах. Парциальные кусты
состоят из небольшого числа (2-3) развивающихся СПФ. Малое их число
лимитируется регулярной затопляемостью местообитания.
Вариант 2. Терминальная почка корневища развертывается в
ортотропный побег, а боковая продолжает нарастать в направлении его роста -
плагиотропно. На следующий год или через 2-3 года терминальная почка вновь дает
ортотропный прирост, на базе которого формируется парциальный куст. Если
отдельные кусты выходят из зоны затопления, то развитие их происходит как у
экобиоморфы 1. Клон образуется рано, через 5-10 лет кусты теряют связь друг
с другом. Во время паводков подземные части побегов обнажаются и разрушаются.
Трансформация биоморф ш. иглистого
Благодаря морфологической пластичности систем побегов,
высокой вегетативной активности, основанной на формировании подземных скелетных
осей, ш. иглистый имеет широкую экологическую амплитуду. Четыре экобиоморфы из разных экотопов составляют
морфологический ряд, в котором наблюдается трансформация форм роста. В ряду
экобиоморф происходят следующие изменения: а) уменьшение размеров растений от
125 до 25 см, б) сокращение длительности основного цикла с 9 до 2-3 лет, в)
сокращение количества СПФ в парциальном кусте от 8 до 2-3, г) уменьшение
продолжительности жизни парциального куста от нескольких десятков до 8-10 лет,
д) изменение в механизмах образования парциальных кустов, приводящее к развитию
низкорослых форм в сочетании с более высокой вегетативной подвижностью, е)
обособление парциальных кустов во все более ранние сроки. Перечисленные
изменения наблюдаются с возрастанием экстремальности существования и именно они
обеспечивают стабильное существование вида даже в крайне неблагоприятной
обстановке.
СПИРЕЯ ИВОЛИСТНАЯ SPIRAEA SALICIFOLIA L.
Спирея иволистная - евразиатский бореальный вид, с основной
частью ареала в умеренной Азии [Гладкова, 1984]. В исследуемом регионе обычна
для территории Северного Охотоморья, верховья Колымы, реже встречается в
Чаунско-Центрально-Чукотском флористическом районе, за исключением его
северо-восточных частей. Приурочена к пойменным лесам, наиболее
предпочтительными местообитаниями являются берега лесных пойменных озер или
влажных заболоченных участков леса.
Материал собран в 1983 г. в верховье р. Кулу по берегам
лесного озера, где спирея образует почти чистые заросли с редкими вкраплениями
отдельных видов кустарниковых ив. Популяция занимает площадь более 2 тыс. м2 и
тянется лентой (7-10 м шириной) по всему периметру озера.
Онтогенез с. иволистной складывается из 4 фаз: 1) проростка (1-й
год жизни), 2) формирования первичного куста (2-7-й годы), 3) образования
парциальных кустов (с 7-го года), 4) развития клона (с 15-20 лет). Фазы
онтогенеза не разграничены четко по годам. Например, формирование первичного
куста может затянуться до 15 лет, а развитие парциальных кустов может начаться
в 5 лет.
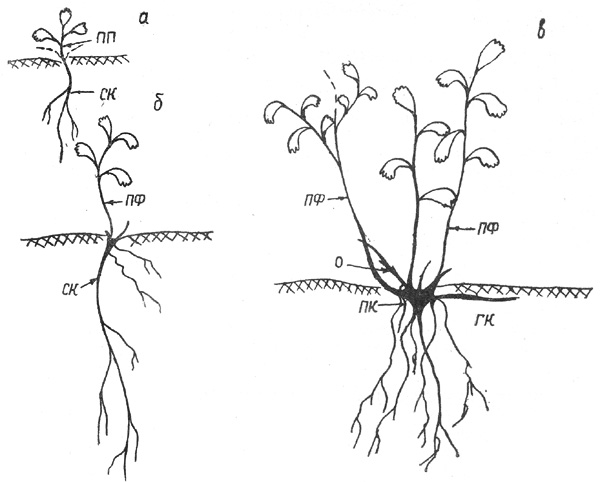
Рис. 6. Спирея иволистная: а - проросток, б - начало
формирования первичного куста, в - первичный куст. Здесь и далее на рисунках О
- отмерший ПФ
|
П е р в а я ф а з а. Проростки встречаются редко, их
трудно отыскать среди огромного числа мелких вегетативных экземпляров. Удалось
найти всего несколько проростков и молодых растений в разреженных зарослях на
некотором удалении (2-3 м) от взрослых плодоносящих кустов. ПП составляет 1-1,2
см; корень, который к концу вегетационного периода разветвляется до 2 порядков
боковых корешков, 3-3,5 см. ПП ортотропный, очень тонкий с 2 семядолями и 3
настоящими листочками (рис. 6, а).
В т о р а я ф а з а: ПП в следующем году не дает нового
прироста, а отмирает почти полностью; лишь основание, скрытое в подстилке (2-3
мм), сохраняется живым и от него отрастает ПФ размером 2,8-3 см. Иногда
одновременно появляются 2-3 ПФ от 1 до 3 см, все они ослабленные и отмирают к
концу сезона почти до основания. От живых частей отрастают ПФ второго порядка
(рис. 6, б) длиной до 5-6 см. В отличие от побегов предыдущего года ПФ второго
порядка отмирают не полностью, а лишь на треть длины. Живая часть побегов в
следующем году акротонно ветвится, образуя 2-3 ПВ, которые также отмирают
верхушкой. Таким образом, возникает СПФ. Более полное отмирание СПФ происходит
к концу 3-го года. Пока идет развитие ПФ второго порядка, от базальной части
куста начинают отрастать новые ПФ (третьего, а затем и четвертого порядков), т.
е. смена ПФ и СПФ в кусте идет не скачкообразно, а постепенно. С возрастанием
порядка ПФ увеличивается продолжительность его жизни и размер прироста 1-го
года, нарастает число СПФ и ПФ в кусте. Основания побегов формируют узел кущения,
который медленно утолщается и в возрасте приблизительно 15 лет имеет диаметр
около 1 см. В узле кущения заложены почки возобновления, которые со временем
реализуются либо в новые ПФ, либо образуют подземные плагиотропные
гипогеогенные корневища. Стержневой корень со временем сильно разветвляется, но
в размерах увеличивается незначительно. На 4-5-й год в районе узла кущения и в
основании побегов появляются придаточные корни. Уже через 2-3 года стержневой
корень становится незаметным среди активно отрастающих придаточных корней.
Корневая система первичного куста напоминает мочковатую. |
Т р е т ь я ф а з а. Первые корневища в узле кущения могут
появиться уже в 5 лет, чаще же это происходит в 12-15 лет. Отдельные почки,
сформированные в узле кущения, после некоторого периода покоя трогаются в рост
и развивают плагиотропный побег (корневище), растущий на глубине 5-12 см, с
приростами 8-18 см в год. На нем закладываются почки, развиваются придаточные
корни (рис. 6, в).
После 2-3-летнего роста корневища некоторые его почки (как
правило, боковые) развивают ортотропный ПФ. Корневище после этого продолжает
нарастать, однако новый прирост дает не терминальная, а ближайшая к ней боковая
почка. То же происходит и в последующие годы. Куртина постепенно разрастается.
На базе появившихся ПФ могут формироваться парциальные кусты. Однако далеко не
каждый ПФ дает начало новому кусту, чаще такие побеги играют роль ПД, выполняя
ассимиляционную функцию. Они слабые, эфемерные, отмирают в год своего
появления.
Дальнейшее развитие ПФ, образующих парциальные кусты,
происходит следующим образом. КО 2-му году ПФ отмирает до уровня подстилающей
поверхности, а живая его часть, скрытая в подстилке или в почве, развивает ПФ
второго порядка. Их приросты больше приростов побегов предыдущего года почти в
3 раза и составляют 13-15 см. В следующем году они замещаются новым поколением
ПФ, которых может быть одновременно 1-3. ПФ третьего порядка отмирают на разную
длину, в отличие от предыдущих ПФ (рис. 7).
Дальнейшее развитие парциального куста мало чем отличается
от материнского, поэтому предлагаемое ниже описание цикла развития побегов и
куста в целом одинаково справедливо для тех и других. Чаще всего ослабленные ПФ
отмирают полностью, т. е. до основания, а более сильные на какую-то часть своей
длины. Эти последние ветвятся, развивая СПФ. Нарастание СПФ в этом случае идет
несколькими путями.
1. Если ПФ отмирает наполовину, то в верхней живой части
отрастают 2-3 ПВ. К концу сезона верхушки этих побегов также отмирают, ниже
образуются ПВ второго порядка, которые в свою очередь также симподиально из
боковых почек образуют ПВ третьего порядка. Эти последние, как правило, в этом
же году дают репродуктивные приросты. ПВ первого-второго порядка имеют размер
5-10 см. Репродуктивный побег приблизительно равен длине вегетативного ПВ
третьего порядка - 9-10 см, он несет 2-4 листа и метельчатое соцветие, слегка
продолговатое по форме с бело-розовыми цветками. Через 2-3 года начинается
отмирание побегов в СПФ, но ей на смену в кусте продолжают нарастать более
молодые по возрасту СПФ и появляться новые ПФ.
2. Если ПФ отмирает только верхней частью, то живая его
часть обильно продуцирует ПВ в числе 10-12. ПВ, расположенные ближе к верхушке,
небольшие - до 1 см, те, что находятся ниже - 10-13 см. Самая нижняя треть
побега не дает ПВ. Спустя 1-2 года здесь могут отрастать эфемерные ПД размером
не более 1 см. СПВ, развивающаяся на ПФ, может иметь до пяти порядков ПВ Все
ПВ, включая первый порядок, дают соцветие. Затем СПФ постепенно деградирует, но
основание его остается живым и отсюда отрастают новые ПФ наряду с теми, что
образуются в узле кущения. Эти дочерние ПФ усиленно растут, достигая размера
45-50 см и, таким образом, их верхушка оказывается на том же уровне или даже
несколько выше уровня побегов отмершей СПФ. В дальнейшем они повторяют цикл
развития материнской СПФ, которую замещают.
3. Иногда на выросшем ПФ усиливаются ПВ первого порядка, причем
если их было 8-1 О и каждый сформировал свою СПВ, несущий их ПФ не выдерживает
нагрузки и полегает. В этом случае в разных частях полегшего побега образуется
несколько ПВ, разных по длине и циклу развития. Слабые побеги располагаются
ближе к верхушке, они недолговечные и вскоре отмирают, более сильные образуются
в основании побега и нарастают в течение нескольких лет. Эти побеги ортотропны,
а отросшие от них ПВ и ПД располагаются под углом к материнскому побегу.
Постепенно прирост 1-го года ПФ достигает максимальной длины
- 50-55 см. СПФ за счет СПВ еще несколько удлиняется, и общая высота куста
достигает 100-110 см. Основной цикл составляет 5-7 лет. Наибольшего расцвета
СПФ достигает к 3-4-летнему возрасту, затем начинается ее постепенное угасание.
Базальная часть побегов входит в состав узла кущения и в этой своей части имеет
большое число почек возобновления, которые ежегодно продуцируют все новые ПФ и
таким образом все время происходит обновление куста. Старые отмирающие и
отмершие СПФ наклоняются и оказываются на периферии куста, а в его середине
идет активное возобновление ПФ. В одном кусте одновременно может развиваться
30-50 ПФ, которые находятся на разных стадиях своего жизненного цикла. |
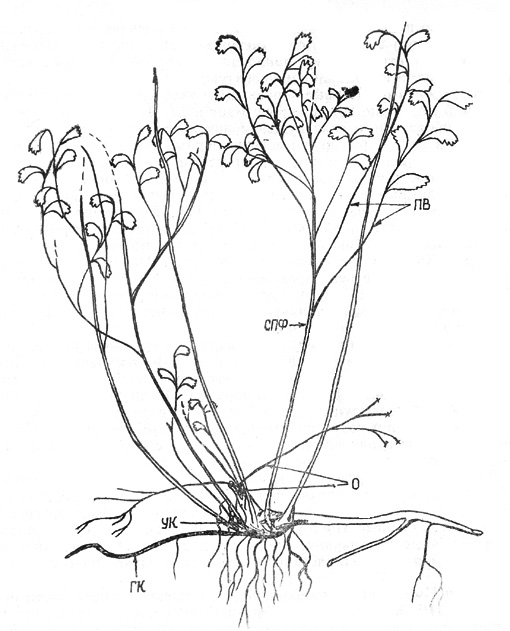
Рис. 7. Спирея иволистная: парциальный куст с отрастающим
корневищем. Здесь и далее на рисунках УК - узел кущения |
Ч е т в е р т а я фаз а. Каждый отдельный куст кроме кроны
формирует и систему подземных корневищ. Сами же корневища, имея большой запас
спящих почек, становятся базой, на которой формируются новые ПФ или ПД.
Появление ПФ означает начало развития парциального куста. ПД изредка образуют
СПД (цикл ее развития не более 2-3 лет), чаще же они эфемерные.
Корневища функционируют не более 10-12 лет, а иногда
разрушаются и раньше. Отдельные парциальные кусты теряют связь друг с другом и
с материнским кустом, продолжая существовать самостоятельно, в результате
цельная куртина распадается, образуется клон. У с. иволистной вегетативное
возобновление преобладает над семенным и часто площадь в несколько десятков
метров может быть занята одним разросшимся клоном. Стареющие кусты в популяции
встречаются редко. В таких кустах число отмирающих и отмерших побегов
преобладает над числом живых и последние все более уменьшаются в размерах.
Репродуктивная активность у них постепенно угасает, на весь куст приходится 3-5
соцветий. Большинство появляющихся ПФ становятся эфемерными, лишь отдельные
образуют СПФ, которая существует 2-3 года, однако размеры такой СПФ значительно
меньше размеров СПФ предыдущих лет. Вскоре куст отмирает. Вычислить
продолжительность жизни отдельного куста, а тем более клона практически
невозможно. С уверенностью можно говорить лишь о том, что куст существует
несколько десятков лет. Что касается клона, то он живет неопределенно долгое
время.
Таким образом, в условиях Верхней Колымы с. иволистная
обладает высокой вегетативной подвижностью. Быстрое разрастание куртины на базе
корневищ и дальнейшее их достаточно быстрое разрушение делает возможным
формирование жизнестойкого клона, способного занимать большие пространства,
вытесняя при этом менее конкурентоспособные виды.
СПИРЕЯ СТЕВЕНА SPIRAEA STEVENII (SCHNEID.) RYDB.
Амфиберингийский гипоаркто-монтанный (подгольцовый
субокеанический) вид, обычный в советской Арктике в пределах гористых районов
крайнего севера-востока Азии, в полосе гипоарктических тундр [Гладкова, 1984].
В исследуемом регионе обычна по всей территории, кроме о-ва Врангеля.
Встречается в лиственничниках, зарослях кедрового стланика, пойменных лесах,
ивняках, ерниках.
Исследования проводились с июня по сентябрь 1983 г. в
верховьях р. Кулу. Наблюдались растения из различных экотопов. Здесь
описывается онтогенез вида, растущего среди ерника, который тянется полосой
более 2 км по приречной террасе р. Кулу. Высота кустов 30-40 см.
В онтогенезе выделено 4 фазы: 1) развитие одноостного побега
(l-2-й годы), 2) начало ветвления и формирование первичного куста (3-7-й годы),
3) формирование парциальных кустов (с 7 до 18-20 лет), 4) образование клона (с
18-20 лет).
П е р в а я ф а з а. Проростки и молодые растения встречаются
на пятнах мелкозема вблизи взрослых плодоносящих кустов. Проросток ортотропный
8-10 мм длиной, с 2-3 настоящими листочками. Семядоли функционируют 2-3 недель,
затем засыхают и обламываются. Стержневой корень 3 см, слабо разветвленный на
верхушке. Размер листьев 2-3 мм. К следующему году ПП отмирает до основания, от
которого отрастает побег текущего года, так же, как и ПП, размером не более 10
мм. По структуре этот побег полностью аналогичен ПП и также отмирает либо до
основания, либо на определенную часть своей длины. Стержневой корень удлиняется
до 5-8 см, разветвляется до 2-3 порядков боковых корешков (рис. 8, а).
В т о р а я ф а з а. На 3-й год на границе отмершей и живой
части возникает новый ПФ, цикл развития которого отличается от побегов
предыдущих лет. К следующему году этот побег отмирает лишь в своей верхушечной
части, живая его часть (3-4 см) обильно ветвится, давая до 6 ПВ, разных по
длине - от 0,5 до 1,5 см. Корень к этому времени сильно удлиняется - до 15-18
см, разветвляется до четвертого-пятого порядков, растет плагиотропно или с
некоторым наклоном к поверхности почвы на глубине 8-10 см. Живые основания
побегов постепенно утолщаются, из них образуется узел кущения, который является
базой дальнейшего формирования первичного куста. Новые ПФ ежегодно отрастают от
узла кущения. Они имеют удлиненные приросты, которые с возрастанием порядка ПФ
все более увеличиваются в размерах; также постепенно происходит удлинение
основного цикла. В первую и вторую фазы онтогенеза смена ПФ идет быстро, так
что к 7-8 годам отрастают ПФ пятого-шестого порядков.
Образующиеся побеги ежегодно отмирают на разную длину.
Отмирание до основания вызывает появление новых ПФ, частичное, активное
ветвление побега. ПВ образуются симподиально. В случае отмирания верхушки
побеги на следующий год дают ПВ второго, затем и более высоких порядков -
формируется СПВ. ПВ второго-третьего порядка примерно на 10-12-й год развития
куста дают репродуктивный прирост из терминальной почки, т. е. за один сезон ПВ
образует два прироста: вегетативный и репродуктивный (1-2 см первый и 5-7 см
второй). Цветонос заканчивается головчатым соцветием с мелкими белыми цветками.
В возрасте 5-6 лет на погруженном в почву основании узла кущения возникают
первые придаточные корни. Одни из них сразу усиливаются иза год достигают 10-12
см, другие остаются тонкими, размером не более 1,5-2 см. Последние отмирают уже
в следующем году, первые становятся многолетними (рис. 8, б). |
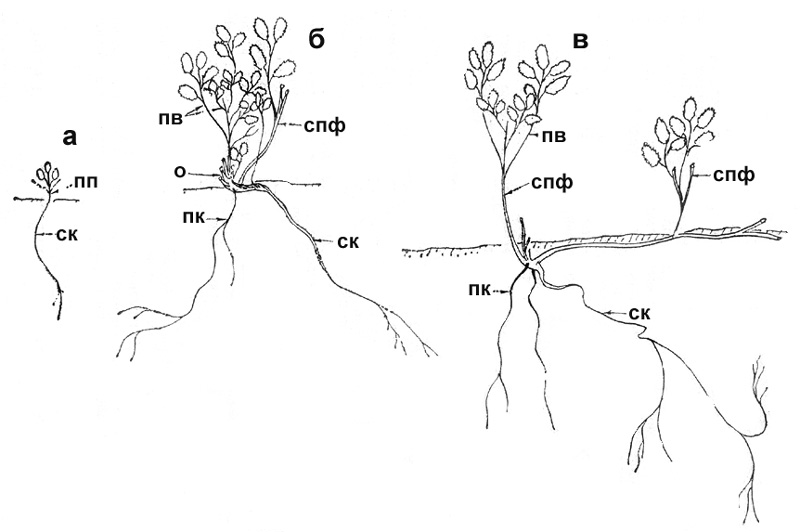
Рис. 8. Спирея Стевена: а – проросток, б – первичный куст, в
– формирующиеся парциальные кусты |
Тр е т ь я ф а з а. В 6-7 -летнем возрасте от погруженной
части узла кущения отрастает плагиотропный побег (гипогеогенное корневище),
вначале небольших размеров - до 1 см, в дальнейшем его приросты значительно
удлиняются. Уже в текущем году корневище может дать ортотропный надземный
побег, который развивается из его боковой почки (рис. 8, в). Размер его около 2
см. В дальнейшем он может сформировать систему симподиальных побегов ветвления.
Если побег отмирает полностью, то образование куста не происходит.
Образовавшаяся СПВ к концу 3-го года отмирает наполовину или большую часть, а в
ее основании развиваются 1-2 ПФ следующего порядка. В дальнейшем формирование
парциального куста идет так же, как у материнского.
Со временем приросты ПФ как в материнском, так и в дочерних
кустах удлиняются и к 13 годам достигают 10-13 см; основной цикл становится
равным 4 годам. ПФ симподиально ветвятся в привершинной части. Некоторые из ПВ
отмирают в этом же году, другие продолжают нарастать и ветвиться, годичные
приросты ПВ в среднем составляют 4-5 см. Однако после 3 лет роста СПФ частично
отмирает, от живой его части активно отрастают ПД. Одновременно их может
появиться от 5 до 7-8. Несколько разные по длине (от 1 до 3,5 см) они могут
быть эфемерными или нарастать 2-3 года, давая уже совсем незначительной величины
приросты - около 1 см. Нарастание всех перечисленных типов побегов не исключает
возможности появления в кусте новых ПФ, напротив, регулярное их отрастание
поддерживает в равновесии систему развивающихся побегов - куст. Каждый ПФ,
пройдя полностью основной цикл, отмирает, а новые нарастают ежегодно, причем
сразу могут развиваться несколько ПФ.
Таким образом, происходит постоянная смена СПФ в растущем
кусте. ПФ развиваются по-разному.
Вариант 1. В год своего появления ПФ стремительно нарастает,
давая такой величины прирост, который позволяет ему достигнуть верхнего яруса
побегов прошлых лет, ветвится такой побег только на следующий год по
симподиальному типу.
Вариант 2. Нарастание и ветвление ПФ идет одновременно,
тогда ПФ достигает несколько меньших размеров, чем в предыдущем случае.
Ветвление побегов здесь, как видим, моноподиальное. Общая высота СПФ в том и
другом случае одинаковая.
Первая, вторая и частично третья фазы соответствуют первому периоду онтогенеза, во время которого происходит
интенсивный рост побегов и накопление биомассы. Наиболее пышными и
продуктивными кусты спиреи становятся тогда, когда СПФ достигают своей
максимальной величины - 30-40 см. Такие кусты насчитывают до 20 растущих ПФ
разного возраста от 1 до 7-8 лет (8-летний основной цикл является наибольшим).
Кусты обильно цветут и плодоносят, в подземной части несут плагиотропные
корневища, на базе которых развиваются все новые парциальные кусты. В таком
состоянии растения вступают во второй период онтогенеза - стабилизации
(нарастание побегов уравновешено их отмиранию) - и находятся в нем длительное
время. Позже наступает третий период, когда постепенно усиливается деструкция
побегов в кусте. Иногда в популяции встречаются стареющие кусты, они выделяются
тем, что число отмерших побегов у них превышает число нарастающих. Отмирание
кустов начинается с того, что новые ПФ перестают появляться ежегодно, к тому же
сокращается длительность их жизни. Меньшими становятся приросты побегов. Со
временем новые ПФ совсем перестают появляться, а старые по завершении цикла
отмирают. В этот период активизируется появление ПД. ОНИ на последнем этапе
жизни куста играют значительную роль, так как отрастают в большом количестве и,
хотя часто они ослабленные и эфемерные, создают дополнительную зеленую массу,
увеличивая тем самым ассимиляционную поверхность куста. Иногда в середине или в
основании отдельных СПФ развиваются ПД с удлиненными приростами, аналогичные
ПФ, но они в сравнении с последними все же значительно меньших размеров, к тому
же очень тонкие и нарастают не более 2 лет, хотя могут ветвиться, образуя СПД.
Таким образом, СПФ постепенно одна за другой отмирают, некоторое время еще
появляются ПД, затем куст отмирает.
Ч е т в е р т а я ф а з а. Корневище, нарастая продолжительное
время в почве, многократно ветвится, развивая сеть подземных побегов, из их
спящих почек формируются парциальные кусты. В течение длительного времени
парциальные кусты с. Стевена сохраняют связь друг с другом, однако со временем
корневища все же разрушаются и дочерние кусты продолжают существовать
самостоятельно. По общим размерам и продолжительности жизни они не уступают
материнскому кусту, каждый является зоной появления новых корневищ, а стало
быть и новых парциальных кустов. Клон разрастается, захватывает все новые
площади, растение переходит к вегегативно-подвижному образу жизни. Определить
границы клона в исследуемой популяции практически невозможно, так как границы
их перекрывают друг друга, выявить принадлежность отдельного парциального куста
к определенному клону не представляется возможным.
ТРАНСФОРМАЦИИ ФОРМ РОСТА В РОДЕ SPIRAEA L.
Род Spiraea в исследуемом регионе представлен 4 видами:
Spiraea salicifolia, S. stevenii, S. betulifolia Pall. (спирея березолистная) и S. media F. Schmidt. (с. средняя). Жизненная форма c.
березолистной ближе к кустарниковой форме, схожей со с. иволистной, а с.
средней - с кустарничком с. Стевена. Спиреи иволистная и березолистная
приурочены в основном к пойменным лесам, долинным ивнякам, где достаточно
плодородный почвенный слой, хорошие условия увлажнения, лучшая защищенность от
разных неблагоприятных воздействий. С. Стевена имеет более широкую
экологическую амплитуду - от пойменных лесов, приречных террас поднимается по
склонам до подгольцового пояса, иногда поселяется на гребнях невысоких сопок на
высоте 800-1000 м над ур. м. С. средняя предпочитает сухие местообитания: опушки
сухих лиственничников, сухие травянистые и степные склоны. Проанализировав эти
данные, можно отметить, что виды рода Спирея образуют морфологический ряд, где
с ухудшением условий существования происходят изменения, ведущие к
трансформации биоморф. Исходными в этом ряду являются вегегативно-подвижные
кустарники с. иволистная и с. березолистная. Они требовательны к эдафическому
фактору, к повышенному увлажнению. С. Стевена более пластична. Она не столь
требовательна к почвенным условиям, к увлажнению, может
поселяться по соседству с вышеназванными видами и тогда
образует кусты до 70 см высотой, формируя биоморфу, близкую к группе
полукустарников. В более суровых условиях приподнятых террас и тем более
склонов и гребней сопок высота кустов 30-50 см, жизненная форма -
вегегативно-подвижный кустарничек. Последним членом ряда является с. средняя,
которая закрепилась и даже предпочитает сухие местообитания с бедными почвами,
сохраняя при этом жизненную форму вегетативно-подвижного кустарничка.
МАЛИНА САХАЛИНСКАЯ RUBUS SACHALINENSIS LEV.
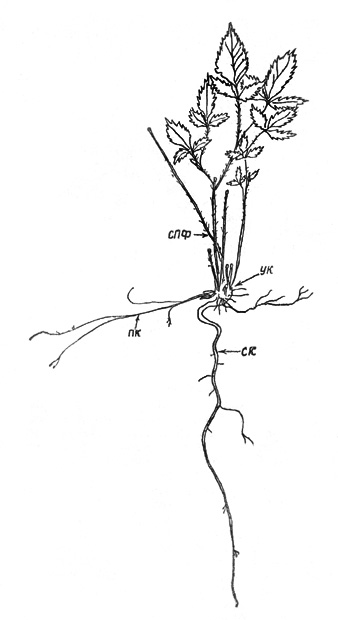
Рис. 9. Малина сахалинская: первичный куст
|
Малина сахалинская - бореальный горнотаежный вид. Ареал его
на территории СССР простирается от бассейна р. Печора через Урал и всю Сибирь
до восточных районов страны - Камчатки, Сахалина, Курил. На юге заходит в
Северную Монголию, Северо-Восточный Китай. В Америке встречается на Юконском
плато, в Североамериканских Кордильерах, равнинных таежных районах Северной
Америки [Гладкова, 1984]. В исследуемом регионе встречается от Прибрежно-Охотского
до Колымского флористических районов по сухим щебнистым и травянистым склонам.
Исследования проводились в 1986 г. в Колымском
флористическом районе, в окрестностях стационара «Контакт». Растет фрагментами
на каменистых россыпях, крупно- и мелкощебнистых осыпях на склонах юго-западной
экспозиции.
М. сахалинская - вегегативно-подвижный кустарник высотой до
130 см. В данном местообитании проростки не найдены, изредка встречаются
молодые вегетативные экземпляры. Начальное развитие растения ввиду отсутствия
проростоков не описано, ниже приводится описание заключительных фаз онтогенеза:
первичного куста, формирования парциальных кустов, образования клона.
П е р в и ч н ы й к у с т. При семенном возобновлении ПП
нарастает, видимо, непродолжительное время, сменяясь отрастающими от его
основания ПФ. Приросты первых ПФ небольшие, о чем свидетельствуют пенечки в
узле кущения. Они тонкие - в диаметре менее 1 мм.
Приблизительно 10-летний первичный куст имеет мощный (30-40
см) стержневой корень, направленный строго геотропно. В районе узла кущения
отрастают придаточные корни, которые, в отличие от стержневого, растут
параллельно поверхности почвы на глубине 10-15 см. Их размер от 3 до 10 см. На
отдельных корнях закладываются почки.
Надземных побегов в кусте 2-3, высотой 10-12 см. Узел
кущения в диаметре около 1 см (0,7-0,8), на нем 7-8 небольших пенечков - оснований
отмерших ПФ. Вычисленный возраст описанных кустиков приблизительно 10 лет.
Первые побеги отмирают ежегодно до основания и только в возрасте 10-12 лет
появляются ПФ с 2-летним циклом. Их прирост В l-й год 10 см, к следующему году
верхушка отмирает, но симподиально отрастают 2-3 ПВ длиной 3-5 см. На 3-й год
СПФ отмирает полностью (рис. 9). С каждым годом приросты ПФ увеличиваются, пока
не достигнут максимальных размеров - 90-100 см. Примерно к 20-25-летнему
возрасту в кусте появляются репродуктивные побеги. Их дают СПФ 2-го года
развития на ПВ текущего года.
Ф о р м и р о в а н и е п а р ц и а л ь н ы х к у с т о в.
Как упоминалось, выше, в узле кущения отрастают придаточные корни, растущие
горизонтально, в год появления они небольших размеров, затем могут достигать
1,5 м и более. На корнях заложены придаточные почки, из которых отрастают
надземные ортотропные побеги. Развитие последних идет следующим образом. Почка
трогается в рост ближе к окончанию вегетационноrо периода (в начале-середине
августа) дает небольшой (1-2 см) ортотропный побег, который в текущем году на
поверхность не выходит (рис. 10). На нем к моменту зимнего покоя формируется
либо только терминальная почка, либо дополнительно 1-2 боковые почки. |
Весной
следующего года (к концу июня) после прогрева почвы терминальная почка
трогается в рост и формирует надземный побег 8-10 см длиной, с 2-3 листьями
(рис. 11). к следующему году побег отмирает до уровня почвы, а из его основания
отрастают новые ПФ. В течение 5-7 лет идет постепенная смена вегетативных ПФ с
годичным основным циклом. Формирующийся узел кущения становится зоной появления
ортотропных ПФ и придаточных корней. Один - из корней растет как стержневой -
геотропно, его основная роль - закрепление (заякоривание) куста, остальные
растут плагиотропно, выполняя функции питания и вегетативного возобновления. На
5-7-й год в парциальном кусте появляются ПФ с 2-летним циклом развития, несущие
репродуктивные органы. В последующем все побеги формирования становятся
дициклическими. Куст постепенно разрастается, увеличивается длина приростов ПФ,
размер узла кущения, число ПФ. Куст существует несколько десятков лет. |

Рис. 10. Малина сахалинская: материнский куст с
горизонтальными корнями и почками на них. Здесь и на рис. 11, 13 П - почки
возобновления |
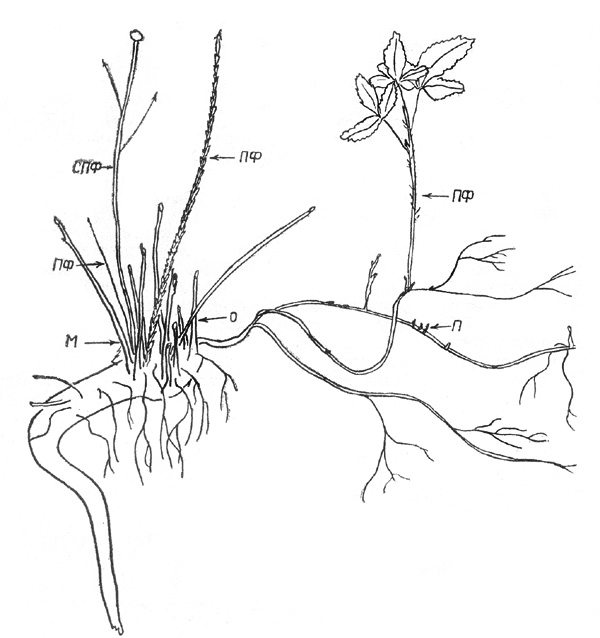
Рис. 11. Малина сахалинская: разросшаяся куртина
|
О б р а з о в а н и е к л о н а. На базе придаточных корней
происходит формирование все новых парциальных кустов, которые с развитием сети
собственных придаточных корней получают все большую автономию. Впоследствии
корень, связывающий куст с материнским растением, разрушается и куст становится
полностью самостоятельным. В дальнейшем он формирует свои дочерние кусты,
способные также обособляться. Так происходит формирование клона, в который
входит большое число куртин, образованных парциальными кустами.
Таким образом, м. сахалинская в данном местообитании
обладает достаточно высокой вегетативной активностью, однако чистых густых зарослей,
как мы наблюдали в районе Охотского побережья, она не образует. Растет
фрагментами среди камней, образуя клон из 10-15 кустов. Малое количество кустов
связано с тем, что здесь неблагоприятные почвенные условия для роста растений
и, кроме того, не каждый ПФ образует новый парциальный куст, часто побеги,
отмирая, не дают замещающих ПФ и выполняют роль ПД, увеличивая ассимиляционную
поверхность куртины. |
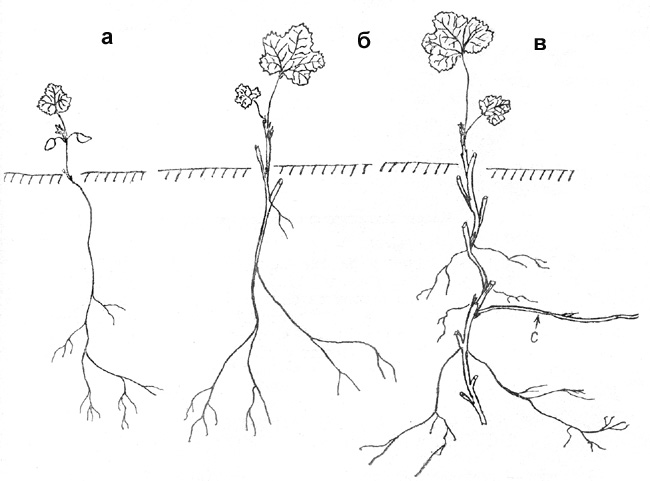
Рис. 12. Морошка: а - проросток. б - симподиально
нарастающий одноостный побег, в - образующиеся столоновидные побеги (С -
столоновидный побег - здесь и на рис. 13, 14)
|
МАЛИНА ПРИЗЕМИСТАЯ, МОРОШКА RUBUS CHAMAEMORUS L.
Морошка - циркумполярный гипоарктическнй вид. На Шпицбергене
и в Гренландии, где известны единичные местонахождения, северный предел ее
распространения [Гладкова, 1984]. На Северо-Востоке СССР обычна по всей
территории, кроме о-ва Врангеля. Поселяется во влажных местообитаниях, часто на
подушках сфагнового мха.
Материал собирался в течение ряда лет в разных районах
Магаданской области. Ниже приводится онтогенез вида, растущего в заболоченной
пойме руч. Контактовый (приток второго порядка р. Кулу) на подушках сфагнового
мха. Исследования проводились в 1986 г.
В онтогенезе выделено 3 фазы: 1) одноостное растение (1-6-й
годы развития), 2) формирование куртины (6-10-й годы), 3) клон (с 10 лет).
П е р в а я ф а з а. Семена прорастают после периода покоя
в начале июля. Семенное возобновление наблюдается редко, основной способ
размножения вегетативный. Проросток (рис. 12, а) размером 1 см, стержневой
корень 4,5-5 см с мелкими волосовидными боковыми корешками. ПП несет 2 овальные
семядоли и 2 листа, один в зачаточном состоянии, развернут не полностью, другой
около 5 мм в диаметре.
В пазухе нижнего листа к концу сезона закладывается почка.
ПП полностью погружен в мох, только листья достигают его поверхности. На
следующий год ПП отмирает, но почка, находящаяся в его основании, дает новый
побег с 1-2 листьями, листовая пластинка более развитого из них около 1 см в
диаметре (рис. 12, б).
В т о р а я ф а з а. Симподиальное нарастание СПП
продолжается в течение ряда лет. Однако на 5-6-й год одна из спящих подземных
почек дает горизонтальный столоновидный побег, который через 2-3 года
приобретает восходяще-ортотропное направление роста, закладывает терминальную
почку и на следующий год выносит на поверхность 1-2 листа, формируя таким
образом дочернюю особь. Так начинается формирование куртины растения. К б-му
году стержневой корень материнского растения отмирает, а СПП с помощью
придаточных корней все более погружается в мох и далее в торф на глубину 10-15
см. |
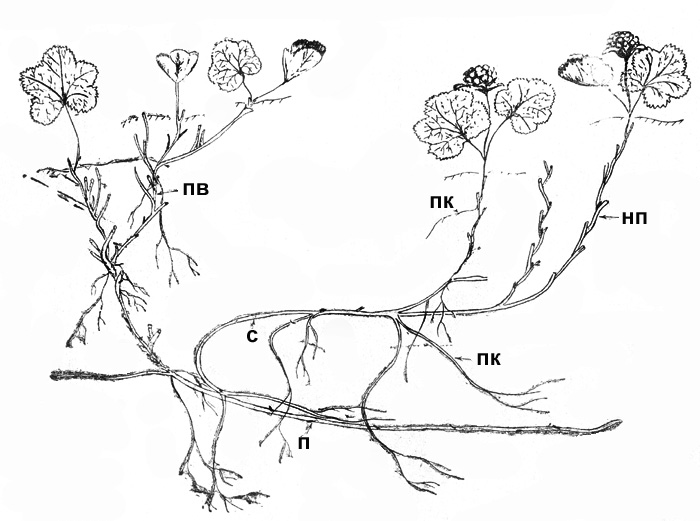
Рис. 13. Морошка: часть клона (НП – надземный побег)
|
Т р е т ь я ф а з а. Некоторые из спящих почек на
столоновидном корневище развертываются в надземные побеги, которые нарастают
одновременно с материнским. В дальнейшем дочерние побеги могут либо отмирать,
либо становиться многолетними, и тогда отрастание новых годичных приростов у
них происходит по симподиальному типу, как у СПП. По-прежнему все системы
побегов находятся в толще моховой подушки, ветвятся, развивая систему
ортотропных побегов, состоящих из небольших по величине симподиальных
приростов. В разных частях погруженных побегов, чаще в их основании,
формируются гипогеогенные корневища (столоновидные побеги), которые также могут
ветвиться, развивать систему корневищ (рис. 12, в). Они несут спящие почки и
придаточные корни, которые чаще всего по 2 отходят от места расположения спящих
почек.
Придаточные корни образуются не только на корневищах,
но и на ортотропных побегах старше 1 года (рис. 13). Со временем ортотропные
надземные побеги на большое расстояние удаляются друг от друга и от
материнского побега, за счет обильно отрастающих корней получают все большую
автономность и в конце концов вследствие разрушения корневищ утрачивают связь
друг с другом. Образуется клон из множества особей.
Как правило, выявить границы клона не представляется
возможным, так как происходит перекрывание одного клона другим, корневища
переплетаются. Популяция занимает пространство в 500-600 м2. Рядом растут виды,
характерные для подобных местообитаний: Andromeda polifolia L., Chamaedaphne calyculata ( L.) Moench., Oxiccocus microcarpus Turcz. ех Rupr.
и некоторые другие. В состоянии клона морошка может существовать неопределенно
долгое время. |
МАЛИНА АРКТИЧЕСКАЯ, КНЯЖЕНИКА RUBUS ARCТICUS L.
Княженика - арктобореальный (низкоарктическо-бореальный)
вид, весьма изменчивый на протяжении своего обширного ареала [Гладкова, 1984).
В исследуемом регионе обычна по всей области, не отмечена лишь на о-ве
Врангеля. Предпочитает пойменные леса, в том числе лиственничники, ерники,
ивняки, ольховники.
Описание биоморфологии вида дано по материалу, собранному в
районе стационара «Контакт» в 1986 г.
Княженика в условиях Северо-Востока СССР возобновляется в
основном вегетативным путем, иногда происходит вызревание семян, но это явление
достаточно редкое. В данном местообитании популяция занимает пространство в
несколько десятков метров, представлены исключительно вегетативные экземпляры;
проростки и ювенильные растения в ней отсутствуют, поэтому ниже дается описание
взрослых растений в фазе клона.
Надземная часть растений представлена кустиками разного
возраста. Отрастание молодых вегетативных побегов происходит с момента
оттаивания грунта и до первых чисел августа. Побеги, тронувшиеся в рост в
середине августа и позже, на поверхность не выходят, дают подземный прирост
0,5-1 см, в таком состоянии уходят в зиму и лишь на следующий год
развертываются в надземный ортотропный побег 1-1,5 см длиной. Он несет 2-3
листа, центральная доля которых не более 3-4 мм, черешки до 3 см. Побеги
следующего года отрастают от живых оснований побегов предыдущих лет,
находящихся в почве. Размеры надземных органов растений постепенно увеличиваются
и не столько сам побег (он чаще всего не более 2 см), сколько черешки и
пластинки листьев; максимальная высота кустиков достигает 10-12 см. Такие
взрослые растения кустятся, развивая побеги ветвления по 3-5 на одном побеге.
Иногда в кустиках сохраняются отмершие, но не опавшие побеги предыдущего года,
хотя чаще бывает, что они обламываются и от них остаются лишь основания,
скрытые в почве.
В подземной части растение образует систему разветвленных
корневищ. Они несут многочисленные придаточные корни, которые образуются
ежегодно и чаще бывают однолетними, хотя часть из них усиливается и может
функционировать в течение нескольких лет. По всей длине корневищ расположены
спящие почки или уже тронувшиеся в рост молодые побеги. После нескольких лет
корневища перегнивают, отдельные части растений обособляются, поэтому популяция
представлена клоном, который состоит из множества самостоятельных систем,
несущих в надземной части разновозрастные кустики, в подземной же они
объединены общим корневищем (рис. 14). |
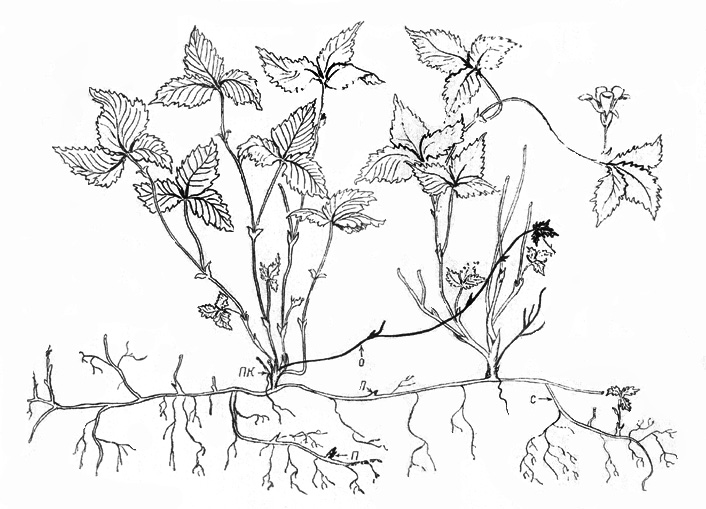
Рис. 14. Княженика: часть клона |
ТРАНСФОРМАЦИЯ ФОРМ РОСТА В РОДЕ RUBUS L.
Три представителя рода Rubus, исследованные нами, относятся
к разным жизненным формам: кустарникам и травянистым многолетникам. Малина
сахалинская - геоксильный кустарник, в надземной сфере образует сменяющие друг
друга СПФ с 2-летним основным циклом.
Развитие куртины начинается примерно в 15--20 лет, клон
образуется в еще более позднее время. Убыстрение смен побеговых систем
надземных побегов, уменьшение их размеров, активизация вегетативной подвижности
приводит в роде Rubus к образованию вегетативно-подвижных травянистых
многолетников, представителями которых являются морошка и княженика. Надземные
побеги у этих растений отмирают ежегодно, а наличие в подземной сфере активно
нарастающих и быстро разрушающихся столоновидных побегов способствует более
быстрому образованию и разрастанию клона. Это, в свою очередь, приводит к
активизации вегетативного возобновления и захвату растениями новых площадей.
Исследованиями И. В. Ивановой [1967, 1968а, б] наглядно показано существование
перехода от кустарников к травам в роде Rubus. Это подтверждается и нашими
данными.
КУРИЛЬСКИЙ ЧАЙ DASIPHORA FRUТICOSA (L.) RYDB.
Азиатско-американский низкоарктическо-бореально-монтанный
вид с островным распространением в горах Западной Европы, Урала и Восточного
Казахстана. В советской Арктике встречается только в ее Восточносибирском и Дальневосточном
секторах. В зарубежной Арктике - на Аляске (восточная часть), в западных
материковых районах арктической Канады [Гладкова, 1984]. В пределах
исследуемого региона распространен повсеместно, но более обычен для южных и
центральных районов, реже встречается в восточных и северных частях.
Материал собран в течение 1977, 1980 гг. в Тенькинском (близ
пос. Кулу), Северо-Эвенском (близ пос. Чайбуха и в верховьях р. Омолон) районах
Магаданской области. Ниже приводится онтогенез вида, растущего на галечнике в
долине р. Жданка (приток верховья Омолона), на высоте 880 м над ур. м. Наряду с
кустарниковыми ивами и березами курильский чай является здесь ценозообразующим
видом.
Онтогенез двух экобиоморф Dasiphora fruticosa (L.) Rydb. (= Роtentilla fruticosa L.), растущих в высокогорьях Алтая и лесостепях
Забайкалья, подробно исследован Л. М. Шафрановой [1964J. Изучение данного вида
в условиях Северо-Востока является логическим продолжением упомянутой работы.
Выводы, сделанные на основании сравнительного анализа биоморф [Шафранова, 1964,
1967J, подтверждаются и нашими исследованиями, но, несмотря на сходство
северной биоморфы с вышеназванными алтайской и забайкальской, между ними
существуют и некоторые различия, о которых речь пойдет ниже.
В онтогенезе северной экобиоморфы выделено пять фаз, которые
совпадают с описанными для забайкальской формы роста [Шафранова, 1964]: 1)
проросток (1-й год жизни), 2) нарастание первичного побега (2-4-й годы), 3)
развитие первичного куста (от начала кущения - 4-5-й год - до образования
дочерних кустов - 30-35-й год), 4) куртина (от 30-35 лет до распадения куртины
на части), 5) клон (от распадения куртины до отмирания).
Пер в а я фаз а. Прорастание семян надземное. Проростки
появляются после периода зимнего покоя к концу июня рядом со взрослыми
растениями. Их легко обнаружить на песчаных отмелях, мелком галечнике,
мелкоземе, в местах, еще не заселенных другой растительностью. В первых числах
июля проросток 3-5 ММ с хорошо развитым и уже разветвленным стержневым корнем
5-5,5 см длиной. На проростке развиваются овальные семядоли величиной 1,5 мм и
2-3 мелких (2 мм) слаборазвитых листочка, рассеченных на 3 доли. К концу
вегетационного периода проросток увеличивается до 7 мм (рис. 15, а).
В т о р а я ф а з а. В течение 2 лет ПП ортотропно
нарастает, увеличиваясь до 1,5-2 см, листья прошлых лет засыхают и
обламываются. В текущем году на ПП развертывается 3 листа размером до 3 мм, на
коротких (2-3 мм) черешках. В основании ПП на приростах 1-2-го года формируются
почки, которые позже реализуются в побеги формирования. Корень разветвляется до
трех порядков. Часто 1-2 боковых корешка растут параллельно поверхности почвы и
по размерам не уступают материнскому корню, длина которого к этому времени
достигает 10 см (рис. 15, б).
Т р е т ь я ф а з а. На 3-4-й год происходит отмирание ПП
почти полностью или частично (отмирает только его верхушка). В зависимости от
этого дальнейшее развитие растений осуществляется разными путями.
Вариант 1. Если отмирает только верхушка ПП, то из пазушных
почек листьев прошлого года развивается 1, реже 2 ПВ, которые замещают
материнский побег. Через 1-2 года более ослабленный побег отмирает, оставшийся
растет еще 2-3 года, после чего замещается новым ПВ - происходит формирование
СПП. На 4-5-й год в базальной части ПП из спящих почек отрастают 3-4 ПФ, иногда
их бывает больше. Начинается формирование первичного куста. Приросты ПВ и ПФ
значительно отличаются друг от друга. ПВ имеют укороченные приросты розеточной
структуры с 2-3 небольшими листочками на коротких черешках сближенных на
верхушках побегов. ПФ в 1-й же год усиленно растут, приросты 2-2,5 см с 4-5
листьями, равномерно по ним распределенными.
Вариант 2. В случае отмирания ПП до основания трогаются в
рост спящие почки, расположенные в живой его части ниже уровня почвы. ПФ,
развертывающиеся на них, усиленно растут и к концу вегетации превосходят по
размерам отмерший ПП. В течение 3-4 лет ПФ нарастает ортотропно, не переходя к
ветвлению, и только потом ветвится, образуя СПФ (рис. 15, в).
Таким образом, по первому варианту идет развитие СПП и СПФ,
по второму - только СПФ. |
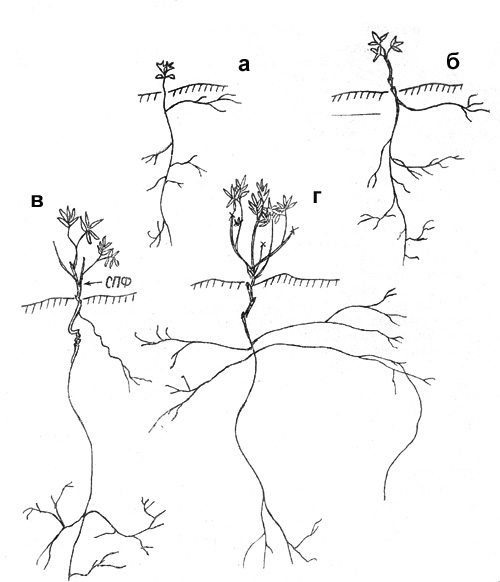
Рис. 15. Курильский чай. Возрастной ряд особей: а -
проросток, б - нарастание ПП (З-й год), в - первичный куст в начале
формирования (8-й год), г - первичный куст (12-й год) |
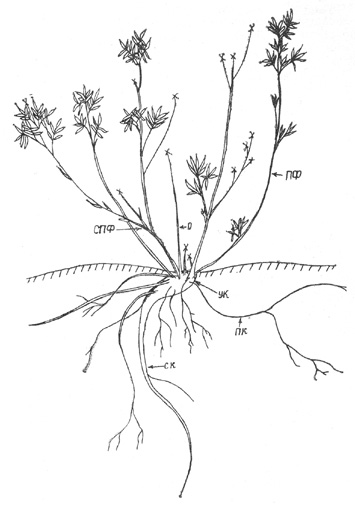
Рис. 16. Курильский чай: материнский куст (15-17 лет)
|
После отмирания ПП образуется первичный куст, формируется
его узел кущения. Б базальной части ПП возникают придаточные корни, а в
основании стержневого корня развиваются многочисленные боковые корешки. Те и
другие способствуют погружению узла кущения в почву (рис. 15, г). Стержневой
корень после нескольких лет вертикального роста изгибается и растет параллельна
поверхности почвы на глубине 10-15 см. СПП и СПФ растут 4-5 лет. В разных
частях их формируются ПД - по 5-6 на побеге, в большинстве они эфемерные,
иногда растут до 2-5 лет. На материнском кусте часто видны пенечки - отмершие
ПД. Для последних характерна небольшая величина годичных побегов (1-2 мм).
Иногда они ветвятся, давая 1-2 также укороченных дочерних побега. Зацветает
курильский чай на 10-12-й годы. Репродуктивные побеги 2-4 см длиной, неразветвленные,
их дают терминальные почки побегов. Растение начинает цвести 7-8 июля, массовое
цветение приходится на середину июля (15-17 июля). К концу вегетационного
периода репродуктивные побеги отмирают, иногда вместе с верхушкой материнского
вегетативного побега, а на следующий год замещаются новыми ПВ. Приросты их
постепенно увеличиваются (максимально до 7 см), они многократно ветвятся.
Большинство образующихся ПБ отмирает в течение 1-5 лет, лишь
2-3 из них нарастают более продолжительное время, становясь скелетными осями
растения. Однако и они по достижении определенного возраста либо частично, либо
полностью отмирают. Отмершие скелетные оси (СПФ) замещаются вновь образующимися
ПФ, которые со временем формируют СПФ, и т. д. Происходит постепенная замена
одних СПФ другими. Длительность жизни каждой новой смены СПФ постепенно
возрастает. Если СПП растет 3-4 года, то СПФ, пришедшая ей на смену, - 7-8 лет,
а следующая - 10-12 лет. Наибольшую длину побегов (до 12 см) образуют ПФ
второго порядка в l-й год.
Узел кущения разрастается, в возрасте 15-17 лет достигает
размера 2,5-3 см, погружается в почву, иногда на значительную глубину - до 10
см (рис. 16). Активность ветвления с возрастом увеличивается, растет и
количество репродуктивных побегов. Многочисленные ПБ развивают цветоносы в год
своего появления, но иногда они нарастают 3-4 года и только после этого дают
репродуктивный прирост.
Достигнув предельного размера (в данном местообитании -
30-40 см, лишь отдельные - до 60 см), растения заканчивают первый период -
накопление биомассы и переходят в следующий - стабилизация развития систем
побегов. В этот период дальнейшего увеличения размера кустов не происходит, так
как отмирание побегов уравновешивается их нарастанием. В кусте одновременно
развивается 20-30 СПФ, обильно ветвящихся в верхней части и находящихся на
разных стадиях своего развития. Растение обильно цветет, по подсчету количества
репродуктивных ПВ на 30 кустах приблизительно одинакового возраста в среднем на
куст их приходится 150-170.
Деградация куста (третий период онтогенеза) наступает не
сразу. Постепенно соотношение нарастания-отмирания побегов смещается в сторону
последнего. Основная масса образующихся ПВ после цветения отмирает, часто
заканчивая цикл развития в 1 год. Уменьшаются их приросты. Появляющиеся в узле
кущения новые ПФ затенены и в связи с этим ослабленные, этиолированные, не
могут по размерам догнать побеги предыдущих лет и после 1-3 лет роста отмирают.
Отмирают и старые СПФ, закончив основной цикл. Узел кущения к этому времени
достигает максимальной величины – 10-12 см в диаметре.
Постепенно число отмерших СПФ нарастает и к концу жизни куста.
остаются 1-2 живые СПФ, остальные 35-45 засыхают, некоторые в таком состоянии
остаются в кусте, другие обламываются. Когда отмирают последние СПФ, жизненный
цикл первичного куста заканчивается. Однако это не конец онтогенеза растения в
целом. |
Ч е т в е р т а я ф а з а. Как упоминалось ранее, на первых
фазах онтогенеза от стержневого корня отрастают горизонтальные боковые корни.
Они возникают и позже. На них развиваются мелкие всасывающие корешки, которые
существуют непродолжительное время. У многолетних корней они сохраняются только
в приверхушечной части. Очень редко горизонтальные корни ветвятся. В момент
отрастания в первичном кусте ПФ третьего-четвертого порядка из придаточных
почек корней развиваются ортотропные надземные побеги. Они нарастают 2-3 года и
отмирают, но из почек, сформированных в их основании (на приросте г-го года),
отрастают ПФ, замещающиеся новыми после нескольких лет роста. В дальнейшем
дочерний куст развивается по типу первичного, но никогда не достигает его
размеров и возраста и отмирает значительно раньше. Приблизительный возраст
наиболее старых из них 20-30 лет. Горизонтальные корни имеют почти на всем
протяжении одинаковый диаметр - от 1-2 мм (молодые) до 3-4 мм (старые) (рис.
17). В месте образования надземных побегов формируется узел кущения, но диаметр
его значительно меньше, чем у первичного куста. Длина горизонтальных корней
может варьировать от 50 см до 1,5 м. Дочерние кусты образуются в разных частях
на расстоянии от 8 до 40-50 см от стержневого корня, отходя либо от его
основания, либо от боковых корней второго и даже третьего порядков. От одного
куста образуется в среднем 3-4, реже 8-12 дочерних.
П я т а я ф а з а. После полного отмирания первичного куста
происходит разрушение основания горизонтального корня, он теряет связь со
стержневым. Происходит распадение куртины. Растение существует в виде клона.
Дочерние кусты имеют размер 15-20 см, иногда больше, они менее долговечны, чем
первичный куст.
Часто продление онтогенеза куста происходит за счет
укоренения полегших стареющих СПФ куста. Полегая, СПФ присыпается почвой и в
этой части укореняется, после чего на СПФ развивается буйная поросль. Чаще в
рост трогаются 4-5 ПФ, а иногда их бывает значительно больше - до 20. В месте
отрастания ПФ происходит утолщение стебля после развития дочернего куста, т. е.
образуется узел кущения. Через 4-5 лет СПФ отмирает, но ей на смену образуются
новые ПФ, которые растут уже более продолжительное время. Иногда дочерние кусты
развиваются настолько быстро, что вскоре достигают высоты материнского. Узел
кущения также не уступает размеру первичного куста. Кроны обоих кустов
смыкаются и образуется как бы единый куст с 2 узлами кущения. Иногда могут
полегать и укореняться 2-3 СПФ, в этом случае формируется куртина, в которой
кусты, как правило, до конца онтогенеза связаны друг с другом. Несмотря на то,
что высота дочерних кустов может достигать высоты материнского, длительность
жизни СПФ и куста все же значительно ему уступает. Определить продолжительность
жизни растения не представляется возможным, так как в течение онтогенеза
происходит неоднократная смена СПФ без какой-либо последовательности. Если
длительность жизни каждой отдельной СПФ можно подсчитать достаточно точно, то
длительность жизни растения в целом можно расценивать лишь приблизительно. Нам
представляется, что она равна примерно 100-150 годам.
Сравнивая онтогенез трех экобиоморф курильского чая из
высокогорий Алтая, лесостепи Забайкалья и северной, можно сделать следующие
выводы. Направления изменений в структуре побегов, отмеченные л. М. Шафрановой
[1964} для забайкальской и алтайской биоморф, получают развитие у северной.
Переход растений в фазу клона у забайкальской формы роста осуществляется на
базе ксилоризом и связан со старением растений. У северной экобиоморфы клон
развивается на основе горизонтальных корней, которые отрастают уже во второй
фазе онтогенеза, а дочерние кусты формируются тогда, когда явных признаков
старения первичного куста нет. Этот факт свидетельствует об усилении
вегетативной подвижности у последнего члена ряда, а также об активном характере
процессов приспособления к экстремальным условиям среды. Вообще активность приспособительных
реакций арктических растений наглядно и на множестве примеров показана А. П.
Хохряковым И М. Т. Мазуренко [Хохряков, Мазуренко, 1983; Мазуренко, 1986] и
полностью подтверждается нашими данными.
Намеченная Л. М. Шафрановой тенденция к геофитизации
надземных побегов у исследованных ею форм находит свое продолжение у
экобиоморфы с верховьев Колымы. У алтайской формы наибольшая высота кустов
80-120 см, у забайкальской в более благоприятных местообитаниях - 70-80 см, в
более суровых - 45-55, у северной экобиоморфы в благоприятной обстановке - 60,
в экстремальной - 35-40 см.
На примере трех биоморф курильского чая из разных
географических точек при движении с юга на север прослеживается тенденция к
трансформации кустарниковой формы роста в кустарничковую на основе таких
приспособительных черт, как усиление вегетативной подвижности, ускорение смен скелетных
осей, миниатюризация побегов и их систем.
|
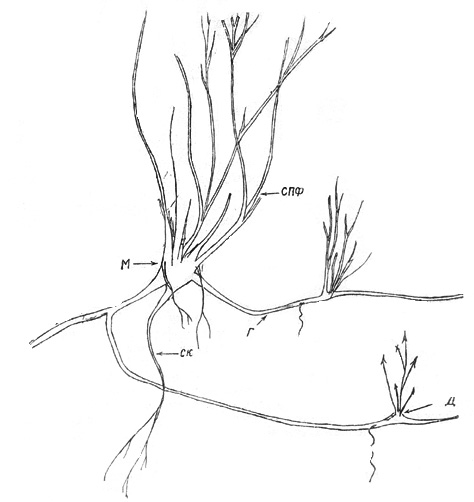
Рис. 17. Курильский чай: разросшаяся куртина (М-здесь и на
рис, 28, 47-материнский куст, Д - здесь и на рис. 28, 46, 47 - дочерний куст, Г
- горизонтальные корни) |
САБЕЛЬНИК БОЛОТНЫЙ COMARUM PALUSTRE
L.
Циркумполярный арктобореальный вид. На территории
Магаданской области распространен повсеместно, кроме о-ва Врангеля. С. болотный
- типичный гигро-гидрофитный вид - встречается по берегам озер, рек, ручьев, в
мелководных осоковых болотах, в сырых лиственничниках. Предпочитает сильно
насыщенные влагой места или растет прямо в воде.
Материал собран в течение четырех полевых сезонов с 1980 по
1983 г. Ниже приводится описание по материалам, собранным в 1983 г. в верховье р. Кулу. На берегу озера, на моховине, наряду с
обильными зарослями взрослых растении обнаружено большое число как проростков,
так и молодых растений. Это дало возможность собрать полный ряд разновозрастных
экземпляров и достаточно подробно изучить и описать онтогенез вида. В данном
местообитании в онтогенезе с. болотного выделено 3 фазы: 1) нарастание ПП
(1-4-й годы жизни), 2) формирование системы ПП (4-10-й годы), 3) существование
отдельных скелетных осей, или фаза клона (от 9-10 лет до отмирания).
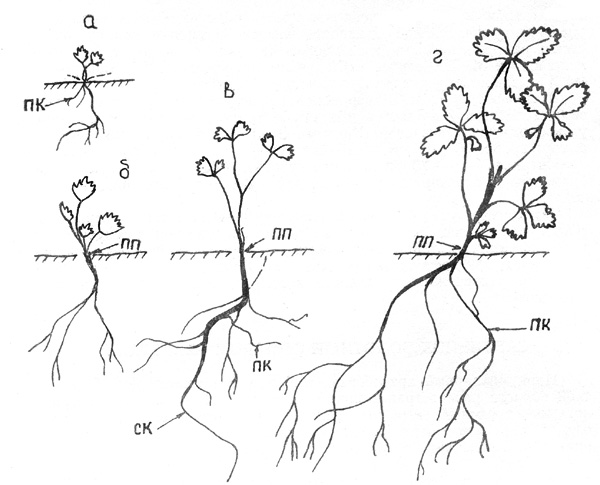
Рис. 18. Сабельник болотный. Возрастной ряд особей: а -
проросток, б - 2-й год, в - З-й год, г - 4-й год
|
Пер в а я ф а з а. Проросток ортотропный длиной 1,5 см. Две
семядоли к концу вегетационного периода, с развитием настоящих листочков,
отмирают и опадают, К концу вегетационного периода развиваются 2, реже 3
округлых не расчлененных на доли листа с мелкими зубцами по краю пластинки.
Размер их от 1 до 3 мм, черешок 3-5 мм длиной. Стержневой корень разветвлен до
двух порядков, размером 3-5 см. Проростки полностью погружены в мох и только
листочки лежат на его поверхности. Придаточные корни развиваются уже на 1-м
году в основании ПП, их 1-3, 3-4 см длиной (рис. 18, а). На 2-й год прирост ПП
составляет 1-1,5 см, размер листьев 8 мм. В основании прироста текущего года
также отрастают придаточные корни. Стержневой и придаточные корни предыдущего
года удлиняются до 10-12 см, разветвляются до четырех порядков. Листовая
пластинка по-прежнему не дифференцирована на доли, но на (каждой пластинке 5
хорошо выраженных зубцов. Прирост несет 4 листа (рис. 18, б). На З-й год особь
увеличивается еще на 2-2,5 см, приросты предыдущих лет погружены в мох и
расположены плагиотропно, а последнего года - ортотропно. Листья на удлиненных
черешках 1,5-1,8 см, пластинка имеет 2 доли, одна покрупнее с 4 зубцами, другая
помельче с З зубцами по краю. Придаточные корни увеличиваются, утолщаются
приросты побегов. Листья предыдущих лет не сохраняются на стебле (рис. 18, в).
В т о р а я ф а з а. На следующий, 4-й год приросты
сохраняются примерно такими же, как в предыдущем году. Происходит дальнейшее
усложнение формы листовой пластинки. Теперь она разделена на 3 равноценные
доли, по краю которых идут зубцы. Самые крупные листья размером более 1 см.
Прирост текущего года ортотропный, предыдущего - восходящий, остальные
расположены горизонтально. Верхушка стержневого корня уже в этом году может
прекратить дальнейший рост, зато боковые и придаточные корни усиливаются. На
молодых частях побегов продолжают активно появляться придаточные корни, в
некоторых частях (чаще всего на стыках годичных побегов) они растут пучком. В
этом году ПП может ветвиться. ПВ один, как правило, ослабленный - всего 5 мм, с
1-2 листьями, на 2-й год обычно отмирает. На приросте ПП 4-5 листьев (рис. 18,
г). В следующие 3-4 года происходит постепенное увеличение размера прироста ПП
до 5-6 см и дальнейшее усложнение формы листовой пластинки. На черешке, ближе к
основанию пластинки листа одна за другой появляются еще 2 доли, теперь листовая
пластинка имеет 5 достаточно равноценных листовых долей. Размер листьев 2-2,5
см, черешков - 3-4 см. Основания черешков расширены и полностью охватывают
стебель. Листья на побеге имеют очередное расположение, междоузлия растянуты.
Предельная величина приростов у сабельника болотного 13-15 см. |
Стержневой корень отмирает в возрасте 7-8 лет, но к этому
времени по всей длине ПП развиты придаточные корни, которые имеют определенный
цикл развития. В 1-2-й годы они небольшие, волосовидные, слабоветвистые, в
следующие 2-3 года усиливаются, достигая максимального размера-17-19 см, разветвляются
до пяти-семи порядков боковых корешков. Но уже 6-7-летние начинают постепенно
деградировать, на 7-9-й год отмирают некоторые из них, на 10-й- остальные,
достигшие этого возраста.
Т р е т ь я ф а з а. После отмирания стержневого корня
начинают отмирать старые части ПП. С 9-10-летнего возраста ПП, продолжая
нарастать верхушкой, отмирает с базального конца. Ветвление ПП становится
теперь более или менее регулярным, но ПВ возникают не каждый год. С возрастом
они усиливаются, пока не достигнут размера ПП. ПВ со временем также переходят к
ветвлению, но, как правило, количество порядков ПВ ограничивается двумя-тремя.
Так как основания побегов постоянно погружаются в субстрат и постепенно
отмирают, то вскоре и отдельные ПВ обособляются и существуют самостоятельно.
Образуется клон. Первые репродуктивные приросты появляются в 7-8 лет, они могут
быть либо одиночными, либо разветвленными, предельная высота их 20-23 см. На 3-4-летних
(иногда более старых) приростах могут появляться ПД. ОНИ более слабые по
сравнению с ПВ, обычно эфемерные, изредка нарастают 5 лет.
Итак, после 8-10 лет растение переходит к
вегетативно-подвижному образу жизни. В дальнейшем в нарастании систем побегов
просматривается определенная цикличность. Годичные приросты достигают
максимального размера - 12-13 см, живая часть побегов имеет возраст не более 10
лет. У отдельных экземпляров отмирание начинается уже в 7 -летнем возрасте,
чаще это происходит тогда, когда основания побегов погружены в воду. Если же
они находятся в почве или мхе, то живут более продолжительное время, но в
данном местообитании все же не более 10 лет. Вновь образующиеся ПВ также
растут, пока не достигнут этого критического возраста, затем основания их
разрушаются, и они отделяются от материнской оси, продолжая существовать
самостоятельно (рис. 19). Постепенно все новые и новые скелетные оси
отчленяются друг от друга, клон разрастается, определить его границы
невозможно. Популяция может занимать площадь 100 м2 и более. Особенно это
характерно для осоковых болот. Большинство озер и берега водотоков в
исследуемом районе окаймлены ковром сабельника болотного по всему периметру.
Основной способ размножения с. болотного в данных условиях вегетативный.
Семенной - редок и, по-видимому, это связано с тем, что проростки и молодые
растения не выдерживают конкуренции быстрорастущего клона.
Таким образом, с. болотный - вегетативно-подвижный
травянистый многолетник, который формируется на основе погружающихся скелетных
осей - эпигеогенных корневищ. Быстрое отмирание корневищ с базального конца
способствует активизации вегетативной подвижности растения. |
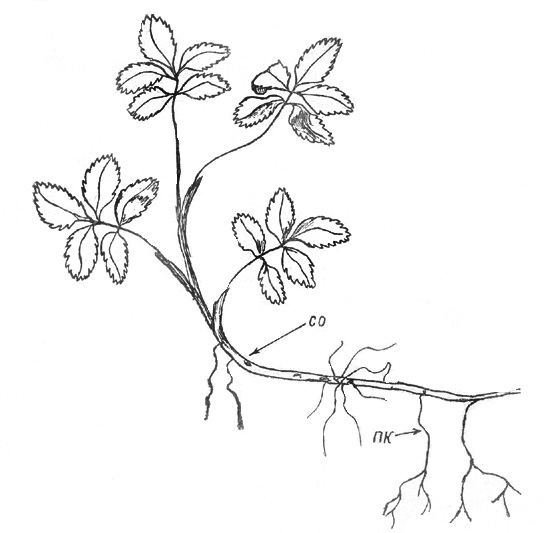
Рис. 19. Сабельник болотный: обособившаяся скелетная ось
(СО) |
|